Ветвление побегов – это образование системы разветвленных осей, благодаря чему увеличивается общая масса надземной части растения [1]. Различают два типа ветвления: верхушечное (дихотомическое) и боковое (происходит за счет моноподиального и симподиального нарастания). При дихотомическом ветвлении рост побега обеспечивается двумя инициалями, образованными в результате деления одной. При моноподиальном способе нарастания верхушечная почка образует главную ось, которая растет из года в год, иногда в течение всей жизни, причем главный стебель, по сравнению с боковыми развивается сильнее. При симподиальном нарастании происходит потеря функциональной активности верхушечной почки, вследствие чего усиливается рост боковых побегов. Форма симподиального ветвления – ложнодихотомическое – апикальная почка отмирает или просто не развивается, а рост продолжают две боковые почки, супротивно расположенные непосредственно под верхушечной почкой [2].
У древесных растений в результате ветвления различного типа образуется крона, то есть совокупность ветвей, побегов и листьев. Биологическое значение кроны состоит в том, что в ней сосредоточена вся листовая поверхность растения, служащая аппаратом фотосинтеза. В ней же располагаются и генеративные органы, то есть цветки и плоды [3]. Форма кроны определяется наследственной природой растительных организмов и отражает приспособление растений к условиям существования.
Функции ветвления и нарастания побеговой системы растения выполняют почки возобновления. Зимой, под покоящимися почками на безлистных ветвях можно увидеть листовой рубец – место прикрепления опавшего листа, а на нём – листовой след – след от оборванных проводящих пучков кроющего листа.
Почка – зачаточный побег. Закономерности заложения листовых зачатков на конусе нарастания побега обусловливают и порядок расположения листьев на стебле. Так как в пазухах листьев находятся почки, из которых развиваются боковые побеги, то от характера листорасположения в большей степени зависит и внешний облик растения. Различают 3 основных типа листорасположения, или филлотаксиса: супротивное, мутовчатое и очередное. Супротивное листорасположение характеризуется наличием в узле 2 листьев – одного напротив другого. При мутовчатом листорасположении от узла отходят более 2 листьев. Наиболее распространено очередное, или спиральное листорасположение, при котором каждый узел имеет только один лист [4].
Стебель – осевой, радиально симметричный орган, обладающий длительным верхушечным (апикальным) ростом, осуществляющий двустороннее передвижение веществ между корнями и листьями, поддерживающий крону растения, способствующий увеличению общей ассимиляционной поверхности растения благодаря ветвлению, участвующий в хранении запасных веществ, а в молодом возрасте – осуществляющий также фотосинтез. Эти функции определяют наличие в стебле покровных, хорошо развитых проводящих, механических тканей и функционально разной паренхимы [4]. Стебель состоит из функционально разных тканей, в расположении которых имеются определенные закономерности [5]. Анализ литературных данных показал слабое освещение знаний по анатомии годичных побегов, поэтому проблема изучения анатомического строения стебля является весьма актуальной.
Материалы и методы исследования
Среди огромного разнообразия деревьев и кустарников в качестве объектов исследования нами были выбраны пять видов древесных растений – липа сердцевидная (Tilia cordata Mill.), черемуха обыкновенная (Padus racemosa L.), тополь черный (Populus nigra L.), сирень обыкновенная (Syringa vulgaris L.), каштан конский (Aesculus hippocastanum L.). Выбор обусловлен широким использованием растений в озеленении города Арзамас (55º37´с.ш., 43º78´ в.д.) и в использовании этих видов в экологических исследованиях, например липа мелколистная относится к дымо- и газоустойчивым породам [6].
В результате исследования биологических особенностей выбранных растений, нами проанализировано и исследовано почкорасположение, листорасположение, тип ветвления, форма кроны, особенности листовых следов, и изучено анатомическое строение стебля годичного побега. Форму листовых рубцов и число листовых следов определяли с помощью бинокуляра МБС – 10. Изучение анатомических особенностей производили на приготовленных лезвием поперечных срезах стебля в области междоузлий с помощью микроскопа Микмед при увеличении 10×20, 10×40. Данные фиксировали в форме фотографий и схем.
Результаты и их обсуждения
Липа мелколистная (Tilia cordata Mill.)
По расположению конечных почек на безлистных ветвях деревьев и кустарников, определили, что для липы мелколистной характерно боковое ветвление, симподиальное нарастание. Настоящая верхушечная почка отпала, оставив маленький рубчик; её место заняла ближайшая к ней пазушная почка. Под пазушной почкой виден рубец на месте опавшего кроющего листа (рис.1). Листовой рубец полукруглой формы, расположенный косо, с одной стороны. Листовой след трехпучковый (рис.2).
У липы мелколистной верхние ветви направлены вверх, боковые – растут почти горизонтально, под прямым углом к стволу, а при старении несколько опускаются вниз. В результате такого анизотропного роста побегов, липе присуща широкоовальная крона. Для побегов липы характерно очередное листорасположение.
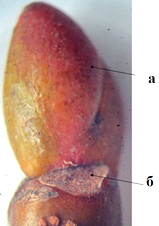
Рисунок 1 – Конечная почка липы (а) с листовым рубцом (б)
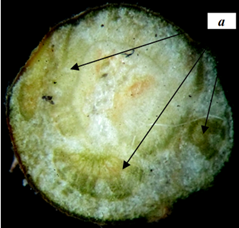
Рисунок 2 – Поперечный срез через трехпучковый (а) листовой рубец липы
На поверхности стебля годичного побега липы мелколистной сохранились остатки отмершей первичной покровной ткани – эпидермы. По мере развития вторичной структуры осевых органов растения эпидерму замещает перидерма. Феллема перидермы 4 – 5-слойная, представлена квадратными клетками (в плане) с утолщенными оболочками. Цвет клеток перидермы придает вызревшему побегу красно-бурую окраску. В перидерме образуются чечевички, через которые осуществляется газообмен живых тканей. Чечевички заполнены рыхлой выполняющей тканью. Под перидермой располагается механическая ткань 4 – 5-слойная пластинчатая колленхима и основная ткань – хлорофиллоносная паренхима (рис. 3). Луб имеет четкую структуру чередования мягкого и твердого в трапециях, разделенных делатационными лучами. В стеблях липы волокна располагаются по периферии флоэмы. Древесина ярусная – горизонтальная слоистость трахеальных и паренхимных вертикальных клеток выдерживается четко. Древесина на поперечном срезе кольца годичного прироста рассеяннопоровая. Сосуды ранней древесины не крупнее или незначительно крупнее, чем сосуды поздней древесины, что согласуется с литературными данными [7]. Для древесины липы характерны, кроме поровых, спиральные сосуды. Распределение осевой древесной паренхимы у липы мелколистной апотрахеальное тяжевое (рис.4), при этом положение паренхимы независимо от положения сосудов, клетки сгруппированы в виде полос, что согласуется с литературными данными [4]. В древесине так же видны радиальные первичные паренхимные лучи, доходящие до сердцевины стебля. Сердцевина стебля имеет очертания окружности и состоит из паренхимных клеток округло-овальной формы. Древесина в годичном побеге липы имеет наибольший объем по сравнению с корой и сердцевиной.
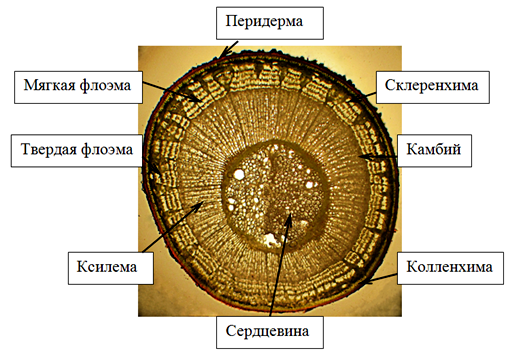
Рисунок 3 – Поперечный срез стебля липы мелколистной
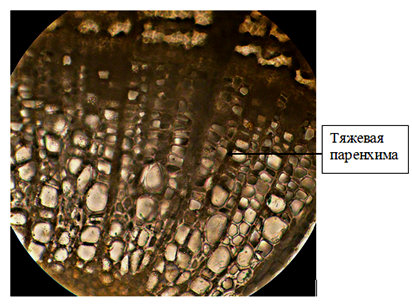
Рисунок 4 – Древесина липы мелколистной (поперечный срез)
Черемуха обыкновенная (Padus racemosa L.)
Выяснили, что главная ось черемухи в течение нескольких лет может нарастать моноподиально за счет верхушечной почки. На 3 – 5 год жизни рост верхушки главной оси прекращается. Под ней образуются из боковых почек две перерастающие её боковые ветви, растущие по направлению главной оси, формируется симподиальное нарастание. Под конечной почкой, являющейся боковой, находиться листовой рубец на месте опавшего кроющего листа, в пазухе которого развивалась эта почка.
У черемухи обыкновенной листовой рубец окружен буро-красноватым валиком (рис.5). Листовой рубец по форме почковидный с трёхпучковым следом. Крона широкояйцевидная. У молодых деревьев ветви отходят вверх под острым углом, с возрастом они пригибаются под действием собственной тяжести книзу. Расположение листьев черемухи обыкновенной очередное.
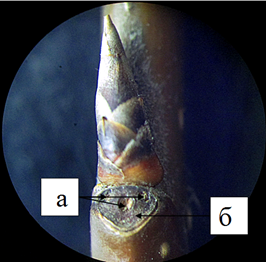
Рисунок 5 – Листовой трехпучковый (а) рубец (б) черемухи
Годичный стебель черемухи обыкновенной покрыт однослойной эпидермой, несущей волоски, которые способствуют защите стебля от сухости или излишней влажности. На смену однослойной первичной покровной ткани эпидермы черемухи обыкновенной приходит перидерма. Феллема перидермы 4 – 5-слойная. Сравнительно тонкостенные клетки феллемы придают вызревшему стеблю черемухи серо-коричневый цвет. Под перидермой находится 3 – 4 слоя сплошной пластинчатой или уголковой колленхимы (рис.6). Флоэма расположена сплошным слоем. Между флоэмой и ксилемой располагается камбий. Древесина черемухи обыкновенной ближе к неярусному виду, поскольку горизонтальные слои трахеальных элементов имеют очень неровные очертания. На поперечном разрезе древесины сосуды распределены равномерно по всему кольцу, но в ранней древесине встречаются отдельные сосуды более крупные, чем в поздней, то есть древесина ближе к рассеяннопоровой. Осевая паренхима представлена апотрахеальным типом, при этом паренхимные клетки сгруппированы в виде полос, но встречаются и рассеянные клетки паренхимы, следовательно, она тяжевого или диффузного вида. На кольце древесины однолетнего побега не предоставляется возможным определить достоверно вид апотрахеальной древесины. Ксилема состоит из крупных и мелких сосудов. Центральный паренхимный цилиндр разделен на две зоны: наружную (перимедулярную) и внутреннюю. По краям наружной зоны выступают 6 следов первичных проводящих пучков древесины. Внутренняя зона состоит из округлых крупных паренхимных клеток.
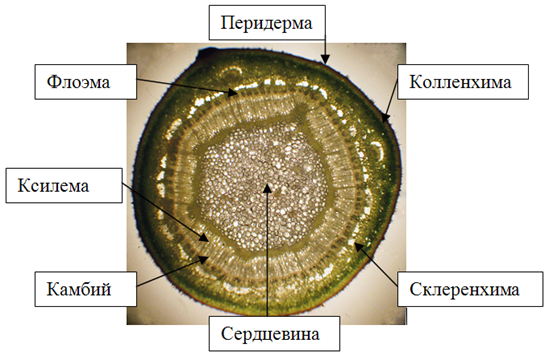
Рисунок 7 – Поперечный срез стебля черемухи обыкновенной
Тополь черный (Populus nigra L.)
Для тополя характерно симподиальное нарастание. Отличительной особенностью тополя черного является чередование удлиненных и укороченных побегов (рис.8). Удлиненный побег характеризуется интенсивным ростом междоузлий. Для укороченного побега характерен заторможенный рост междоузлий, его ось практически состоит из одних узлов, вплотную сдвинутых. Развитием укороченных побегов достигается густота кроны и при занятии сравнительно небольшой площади очень полное покрытие её листвой. Крона – яйцевидная, широкая и раскидистая.
У тополя трёхпучковый листовой след. Пучки листового следа попарно соединены. Листовой рубец сердцевидной формы, либо в виде полумесяца (рис.9). У тополя крупные ветви обращены под углом вверх. Установили, что тополю черному свойственно очередное листорасположение.

Рисунок 8 – Укороченные (а) и удлиненные (б) побеги тополя

Рисунок 9 – Листовой рубец (а) тополя черного с 3 пучками (б)
При вторичных изменениях в стебле тополя черного к концу первого года жизни формируется вторичная покровная ткань перидерма. Её основная часть пробка состоит из двух рядов клеток. Под покровной тканью располагается многослойная первичная кора, состоящая в основном из паренхимных клеток. В паренхиме располагается механическая ткань – пластинчатая колленхима. Склеренхимные волокна перециклического происхождения расположены сплошным кольцом.
Вслед за ними располагаются проводящие ткани флоэма и ксилема, разграниченные между собой камбием. Структура древесины на тангенциальном срезе неярусная. Горизонтальные слои имеют очень неровные очертания. Древесина годичного побега тополя рассеяннососудистая – с более или менее однообразными по размеру и равномерно распределенными по всему кольцу сосудами, что согласуется с литературными данными [1]. Осевая паренхима паратрахеальная скудновазицентрическая, что согласуется с литературными данными [8]. Клетки паренхимы образуют вокруг сосудов сплошную или прерывистую обкладку. Отличительной особенностью клеток сердцевины является их мелкий размер и пятиугольное расположение (рис. 10).
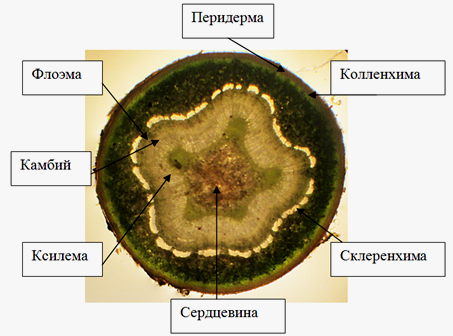
Рисунок 10 – Поперечный срез стебля тополя черного
Сирень обыкновенная
(Syringa vulgaris L.)
У сирени выражена особая форма симподиального нарастания – ложнодихотомическая. При таком типе ветвления верхушечная почка отмирает или не развивается. Рост и ветвление осей в побеговой системе сирени происходит за счет двух боковых, конечных супротивно расположенных почек. Одна из них обычно крупнее, имеет большую емкость. Из неё развивается более сильный побег, который часто по направлению роста продолжает материнскую ось. Крона сирени яйцевидная.
У сирени обыкновенной листовой след пятипучковый, иногда больше. Пучки расположены скученно в виде полумесяца. Листовые рубцы двух супротивных почек не соприкасаются друг с другом (рис. 11). Листорасположение сирени обыкновенной накрестсупротивное.
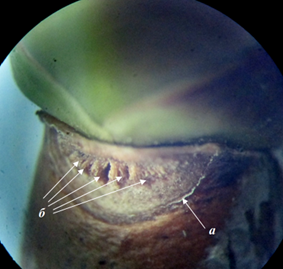
Рисунок 11 – Листовой рубец (а) сирени обыкновенной с 5 пучками (б)
На поперечном срезе годичного побега сирени обыкновенной под омертвевшей эпидермой, представленной одним слоем клеток, располагается перидерма, которая приходит на смену первичным поверхностным тканям. Феллема развита слабо и представлена одним слоем со сравнительно тонкими стенками. В перидерме формируются чечевички, заполненные выполняющей тканью. Под слоем перидермы располагается слой механической ткани – пластинчатой колленхимы. Она представлена четырьмя слоями прямоугольных клеток. Под колленхимой располагается основная паренхима. Склеренхима перицикличексого происхождения, расположена тяжами. После механических тканей располагаются проводящие ткани – флоэма и ксилема, отграниченные друг от друга образовательной тканью – камбием. Структура древесины похожа на ярусную. Древесина ближе к рассеяннопоровому типу – сосуды разбросаны более или менее равномерно в толще слоя прироста, размер и количество просветов только незначительно уменьшается по направлению к внешней границе годичного слоя. Клетки осевой паренхимы сгруппированы в виде полос. Такой тип распределения паренхимных клеток называется апотрахеальным тяжевым. Внутрь от проводящих тканей располагается сердцевина, представленная округлыми паренхимными клетками (рис.12).
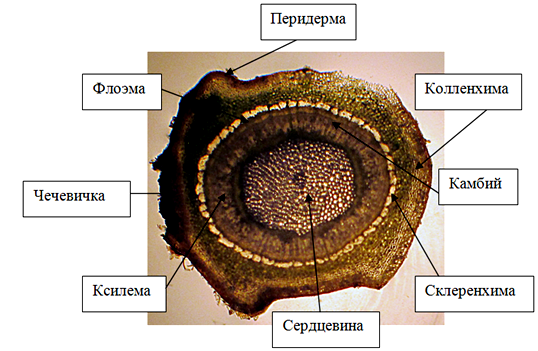
Рисунок 12 – Поперечный срез стебля сирени обыкновенной
Каштан конский (Aesculus hippocastanum L.)
Тип ветвления побегов у каштана конского, как и у сирени – ложнодихотомический. Крона каштана конского – шаровидная. У каштана конского листовой рубец щитообразный, у верхушечной почки трехпучковый, а у боковых – пятипучковый (рис.13). Побеги каштана конского имеют накрестсупротивное листорасположение.

Рис.13. Листовой рубец (а) каштана конского с 5 рубцами (б)
Снаружи стебля каштана конского располагается вторичная покровная ткань перидерма, в которой хорошо развита пробка (феллема), состоящая из 5 уплощенных слоев красноватых толстостенных клеток, придающих стеблю соответствующую окраску. Клетки феллемы прилегают друг к другу плотно, без межклетников. Под перидермой располагается первичная кора, состоящая из механических тканей: пластинчатой колленхимы и ассимиляционной паренхимы. В центральном осевом цилиндре тяжи склеренхимы чередуются с паренхимой. Далее располагается вторичная флоэма (луб). На границе луба и древесина находится камбий. Древесина ярусная – горизонтальная слоистость выдерживается четко, что согласуется с литературными данными [7].
В ранней древесине сосуды более крупные, тип древесины по распределению сосудов – кольцесосудистый, что согласуется с литературными данными [1]. Апотрахеальная тяжевая древесина с элементами рассеянной. В ксилеме выражены спиральные сосуды наряду с другими типами. Центр стебля занят сердцевиной. Она состоит из паренхимных округлых клеток (рис.14).

Рисунок 14 – Поперечный срез стебля каштана конского
В результате проведенных наблюдений, установили, что липа мелколистная и тополь черный имеют симподиальный тип ветвления. Черемуха обыкновенная сначала нарастает моноподиально, а затем симподиально. Растения, характеризующиеся накрестсупротивным типом листорасположения (сирень обыкновенная и каштан конский) имеют ложнодихотомическое ветвление.
Особенности листовых рубцов и листовых следов различны у разных видов древесных пород, что определяется систематикой каждого вида.
В результате проведенных наблюдений, выяснили, что анатомия годичного побега исследованных видов древесных растений имеет видовые особенности, связанные с систематическим положением и общие черты строения, обусловленные жизненной формой растения и возрастом побегов.
В эволюционном аспекте ярусную древесину можно считать более высокоспециализированной, чем неярусную. Ярусная древесина характерна для липы, сирени и каштана. Кольцепоровый тип древесины, по-видимому, является высокоспециализированным и встречается у сравнительно немногих видов, большинство из которых относится к обитателям северной умеренной зоны. Проведение воды в кольцепоровой древесине осуществляется примерно в 10 раз быстрее, чем в рассеяннопоровой [7]. Из пяти видов древесных растений в стебле однолетнего побега каштана конского древесина напоминает кольцепоровую по типу расположения и размеру диаметра сосудов ранней и поздней древесины, а у всех остальных растений – рассеяннопоровую.
Библиографический список
- Андреева И. И., Родман Л. С. Ботаника. – М.: Колос, 2002. – 488 с.
- Широкова Н.П. Особенности строения стеблей и листьев некоторых видов растений // Современные тенденции в образовании и науке: сборник научных трудов по материалам Международной научно-практической конференции 31 октября 2013 г. : в 26 частях. Часть 4; М-во обр. и науки РФ. Тамбов: Изд-во ТРОО "Бизнес-Наука- Общество", 2013. С. 150-153.
- Федоров А. А. Атлас по описательной морфологии высших растений / А.А. Федоров, М. Э. Кирпичников, З. Т. Артюшенко – Ленинград: Издательство Академии Наук СССР, 1956. – 301 с.
- Лотова Л. И. Анатомия и морфология высших растений – М.: Эдиториал УРСС, 2001. – 528 с.
- Широкова Н. П., Недосеко О. И. Избранные темы анатомии и морфологии растений: учеб. Пособие – 3-е изд., перераб. и доп. – Арзамас: АГПИ, 2012. – 169 с.
- Бухарина И. Л., Двоеглазова А. А. Биоэкологические особенности травянистых и древесных растений в городских насаждениях. – Ижевск: Удмуртский университет, 2010. – 184 с.
- Эзау К. Анатомия семенных растений. Т. 1. – М.: Мир, 1980. – 218 с.
- Яценко-Хмелевский А.А. Основы и методы анатомического исследования древесины. – М. – Ленинград: Издательство Академии Наук СССР, 1954. – 338 с.