Electrophysiologiucal investigation of the action of the low doses of thyroxine on background electrical activity of single pyramidal neurons of rat’s cortex under left side lateral hemisection of spinal cord.
In these series of investigations the question of the use of low doses of thyroxine on rats in norm and under left side lateral hemisection of rat’s spinal cord is discussed. The obtained results show the protective effect of thyroxine on extracellular background electrical activity of single pyramidal neurons of rats with left side lateral hemisection. The registration and analysis of the background electric activity of single pyramidal neurons of rat’s cortex is done by means of the special computer programs in on-line mode.
T. S. Khachatryan
Institute of Applied problems of Physics of NAS RA, Yerevan
Общеизвестна роль тиреоидных гормонов в обеспечении нормального развития нервной системы живого организма. В литературе имеются данные относительно применения экзогенных тиреоидных гормонов, приводящих к улучшению функционального восстановления после травмы периферической и центральной нервной системы [1]. Другие научные исследования свидетельствуют о небольшом улучшении функционального восстановления в повреждённом спинном мозге (СМ) крыс при действии экзогенного трийодтиронина в течение двух недель после спинномозгового повреждения [2, 3]. На животных с латеральной гемисекцией (ЛЛГМС) СМ и действием тироксина (Т4) показано быстрое восстановление соматических функций, сопровождающееся более выраженной гипертрофией нейронов и разрастанием нервных волокон, а также наличием слабо выраженных дистрофических и рубцовых изменений в повреждённой области СМ [4].
В наших предыдущих исследованиях, была подробно изучена роль Т4 в изменении электрической активности интернейронов и мотонейронов повреждённого СМ крыс и был представлен подробный анализ данных, касающихся влияния Т4 на изменение электрической активности нейронов СМ крыс в норме и при патологии [5] .
Нашими исследованиями на крысах, подверженных ЛЛГМС и дествию Т4 с регистрацией электрической активности спинномозговых потенциалов были получены предварительные данные указывающие на активирующее действие гормона щитовидной железы (ЩЖ) [6, 7]. В данном исследовании представлены данные, касающиеся влияния Т4 на изменение внеклеточной фоновой (ФА) и вызванной (ВА) электрической активности одиночных пирамидных нейронов (ПН) коры больших полушарий (КБП) головного мозга (ГМ) крыс при органических повреждениях СМ (ЛЛГМС).
Материал и методы
В данной серии научных исследований опыты ставились на 60 белых крысах – самцах, массой (200 – 220 г), разделённых на 4 подопытные группы: первая – 10 экземпляров – интактные животные; вторая – 10 – экземпляров – животные с ЛЛГМС СМ на уровне Т8 – Т9; третья – 20 экземпляров – животные, без спинномозговых повреждений, получавшие в течение двух недель после операции ежедневно инъекции гормона ЩЖ Т4 (дозировка – 50 мкг/кг массы животного); четвёртая – 20 экземпляров – животные с ЛЛГМС СМ, поизведённой на вышеотмеченном уровне, получавшие в течение двух недель после операции ежедневно инъекции гормона ЩЖ Т4 (дозировка – 50 мкг/кг массы животного).
Животные оперировались в стерильных условиях под внутрибрюшинном нембутало – хлоралозовым наркозом (30 мг/кг массы тела и 15 мг/кг массы тела). Под брюхо наркотизированного животного подкладывался небольшой валик и передние лапы подтягивались к задним. В грудной части спины выстригали и выбривали шерсть, затем смазывали кожу йодом. Операционное поле обкладывалось стерильными салфетками, после чего производили разрез по линии остистых отростков на уровне грудного отдела позвоночника. Мышцы по сторонам отростков отслаивались в стороны до самых дужек позвонков. Маленькими костными щипцами резецировались остистые отростки двух позвонков, специальными крючками раздвигались дужки позвонков. С помощью утончённого катарактального скальпеля перерезали спинной мозг и его оболочки по окружности на уровне Т8 – Т9. После этого полноту перерезки проверяли осторожным приподнятием концов мозга и обследованием их при орошении операционного поля слабой струёй физиологического раствора, а также натяжением хвоста животного. Затем приостанавливали кровотечение с помощью местного кровоостанавливающего средства с последующим тщательным орошением физиологическим раствором. После этого узловыми швами сшивали мышцы и кожу.
По завершении данного срока на животных были поставлены электрофизиологические эксперименты. Микроэлектрофизиологическими методами с применением стеклянных микроэлектродов (диаметр кончика 2 – 4 микрон, заполненных 2М раствором NaCl) производилась запись внеклеточной ФА корковых ПН I – IV слоя КБП ГМ крыс. Анализ ФА одиночных ПН производили посредством специальных программ на компьютере (on-line). Выборка спайков проводилась с помощью амплитудного дискриминатора посредством программного анализа. Вычислялась скользящая частота (для каждых 10 межимпульсных интервалов с шагом 5 интервалов), сериальные гистограммы до 50 порядка, гистограммы межимпульсных интервалов на основе которых вычислялась средняя частота фонового нейронального потока одиночных клеток. Затем, по вышеотмеченной методике, производили экстраклеточную регистрацию ФА одиночного ПН КБП ГМ с I по IV слой КБП ГМ. Более подробно с методикой проведения наших экспериментов можно ознакомиться в наших предыдущих публикациях [8, 9].
Результаты и обсуждение
На рисунке 1 представлена картина внеклеточной ФА одиночных ПН в I – IV слоях КБП ГМ крыс у интактных животных. Анализируя данный рисунок, можно сделать вывод, что при послойном отведении внеклеточной ФА одиночных ПН I – IV cлоёв КБП ГМ у интактных крыс, регистрируется регулярный тип разряда данных нейронов, наиболее выраженный в I, II слоях КБП. На рисунке 2 представлены данные регистрации ФА одиночных ПН в I – IV слоях КБП ГМ у животных c органическими повреждениями СМ типа ЛЛГМС. На данном рисунке наглядно иллюстрируется эффект угнетения ФА одиночных ПН КБП ГМ крыс, возникший, вероятно, в результате органического повреждения СМ, наиболее выраженный в I – III слоях КБП ГМ крыс по сравнению с интактными животными (рис.1). Для данных нейронов характерен «пачечный» патологический тип разряда, ранее исследованный нами на интернейронах и мотонейронах СМ у крыс с ЛЛГМС и правосторонними латеральными гемисекциями СМ, а также и при более тяжёлых патологиях СМ, типа 2/3 перерезки СМ и полных хордотомий.
На рисунке 3 представлена картина ФА оди¬ночных ПН в I – IV слоях КБП ГМ у животных без спинномозговых повреждений, получавших систематические инъекции тиреоидного гормона Т4 в течение двух недель по вышеуказанной дозировке. Анализируя данный рисунок можно сделать вывод о том, что при систематическом введении Т4 с последующей регистрацией ФА одиночных ПН КБП ГМ, наблюдается картина электрофизиологических показателей в целом близкая к таковой у интактных животных. Однако, в картине ФА одиночных ПН КБП ГМ при действии Т4 нами наблюдались также некоторые отличия от вышеуказанной активности у интактных животных, основное из которых заключалось в резком учащении частоты фонового нейронального потока, заключающимся, вероятно, в стимулирующим ФА действием данного тиреоидного гормона.
На рисунке 4 представлен эффект нормализации внеклеточной ФА одиночных ПН I – IV cлоёв КБП ГМ у крыс с ЛЛГМС СМ, после двухнедельного систематического введения гормона Т4 в место повреждения СМ. В записи внеклеточной ФА одиночных ПН I – IV cлоёв КБП ГМ у крыс с органическими повреждениями СМ мы наблюдали следующий феномен – у животных, получавших Т4 картина ФА одиночных ПН I – IV cлоёв КБП ГМ по своим электрофизиологическим показателям приближается к таковой у интактных животных. Это свидетельствует, вероятно, о стойких процессах восстановления у животных, получавших ежедневные систематические инъекции гормона ЩЖ – Т4.
Эффекты тиреоидных гормонов на нервную систему можно рассматривать в различных аспектах, что связано, с одной стороны, с многообразием структурных уровней нервной системы, а с другой – с многообразием функций, выполняемых нервной системой в организме животных. Известно, что у крысят после удаления ЩЖ сразу же после рождения, вес мозга заметно ниже, чем в контроле; размеры нервных клеток уменьшаются главным образом за счёт цитоплазмы. Число дендритов и аксонов, а также длина отростков резко уменьшаются, а также и плотность их сплетений. Наиболее резкие изменения происходят в IV слое КБП ГМ. Влияние тиреоидных гормонов на ПН КБП ГМ связано в первую очередь с изменениями в сопротивлении мембран этих клеток и изменениями их метаболизма. Нарушения этих двух звеньев могут, вероятно, повлечь за собой цепь нарушений в деятельности нервной системы, явными проявлениями которых являются такие патологии, как гипотиреоз и гипертиреоз [5, 10].
Изучение процессов регенерации у млекопитающих приняло систематический и планомерный характер после того, как наметились перспективы выявления основных условий, стимулирующих, или, напротив, подавляющих регенерационную способность в различных органах и тканях. В настоящее время трудно назвать какой-нибудь орган или ткань у млекопитающих животных, не проявляющие в определённых условиях способности к регенерации. Довольно обстоятельно изучено влияние гормональных начал на процессы роста и дифференцировки молодого организма и разнообразные явления пролиферации составляющие основу физиологической и репаративной регенерации. Накопившееся исследования дают основания для некоторых обобщений, касающихся как физиологического действия гормонов,так и специфичности их влияния на восстановительные процессы. В арсенале современной гормонтерапии значительный удельный вес принадлежит гормонам ЩЖ и кортикостероидам [11].
Проведенные нами исследования, показали, что у животных с органическими повреждениями СМ, типа ЛЛГМС СМ, получавшим каждодневные инъекции малых доз гормона ЩЖ – Т4 в место повреждения СМ, наблюдается наиболее стойкая картина восстановления внеклеточной ФА ПН. У крыс со спинальными повреждениями вышеуказанного типа не получавших инъекций Т4, картина восстановления данных электрофизиологических показателей выражена значительно хуже, либо, в некоторых случаях, полностью отсутствует. На основании проведенных исследований можно сделать вывод о целесообразности применения гормона ЩЖ Т4 при органическом повреждении СМ, типа ЛЛГМС.
Литература
1. Cook R. A., Kiernan J. A. Effects of triiodthyronine on protein synthesis in regenerating peripheral neurons. 1976, J. Exp. Neurol., № 52, pp. 515 – 524.
2. Tator C. H., Van der Jagt R. A. C. The effect of exogenous thyroid hormones on functional recovery of the rat after acute spinal cord compression injury. 1980, J. Neurosurgical, № 53, pp. 381 – 384.
3. Tator C. H., Rivlin A. S., Lewis A. J., Schmoll B. The effect of triiodo – L – thyronine on axonal regeneration in the rat spinal cord after acute spinal cord injury. 1983, J. Neurosurgical, № 58, pp. 406 – 409.
4. Матинян Л. А., Аллавердян А. Г. Роль тиреоидных гормонов в состоянии пластичности у грызунов при повреждении спинного мозга. 1978, В сб.: 7 – ое научное совещание по эволюционной физиологии, посвящённое памяти академика Л. А. Орбели, с. 154 – 155.
5. Хачатрян Т. С., Матинян Л. А., Андреасян А. С., Киприян Т. К. Роль тироксина в изменении электрической активности интернейронов и мотонейронов повреждённого спинного мозга крыс. Ж. Вопросы теоретической и клинической медицины, 2002, № 1, с . 40 – 45.
6. Хачатрян Т. С. Действие лидазы и тироксина на фоновую электрическую активность одиночных пирамидных нейронов коры больших полушарий крыс. Ж. Биолог. Ж. Армении, 2007, № 3 – 4, т. LIX, c. 198 – 202.
7. Хачатрян Т. С. Протекция тироксином изменений вызванной активности повреждённых травмой одиночных мотонейронов спинного мозга крыс. Ж. Биолог. журн. Армении, 2008, № 3,т. LX, с. 64 – 67.
8. Киприян Т. К., Матинян Л. А., Хачатрян Т. С., Чавушян В. А. Электрическая активность нейронов спинного мозга крыс при действии тироксина, гипертиреозе и гипотиреозе. Ж. Вопросы теоретической и клинической медицины, 1999, т. 2, № 7 (14), с. 50 – 54.
9. Хачатрян Т. С., Нагапетян Х. О., Матинян Л. А. Фитотерапия в сочетании с тироксином при органических повреждениях спинного мозга. Ж. Биолог. журн. Армении, 2008, № 4,т. LX, с. 51 – 55.
10. Eyars G., Horn S. Hormones and subcellular structures. 1956, USA, SCIENCE, v.25,pp. 15 – 23.
11. Войткевич А. А. Восстановительные процессы и гормоны. Изд. «Медицина», Л., 1965, с. 3 – 9.
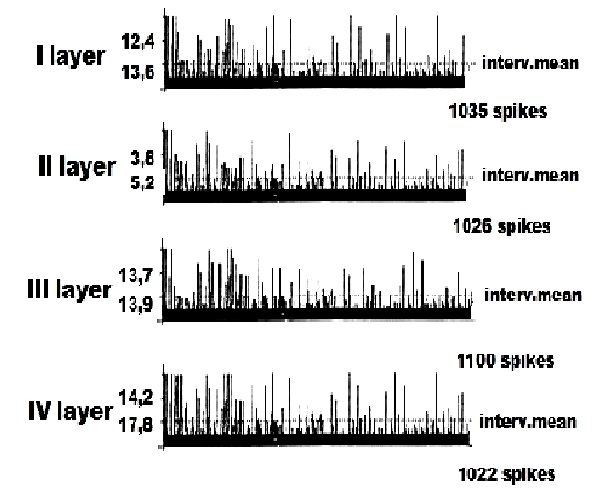
Рисунок 1. Пример программного анализа внеклеточной фоновой электрической активности одиночных пирамидных нейронов I – IV cлоёв коры больших полушарий головного мозга у крыс в норме. Представлена частота импульсов в реальном времени и средняя величина межимпульсных интервалов.
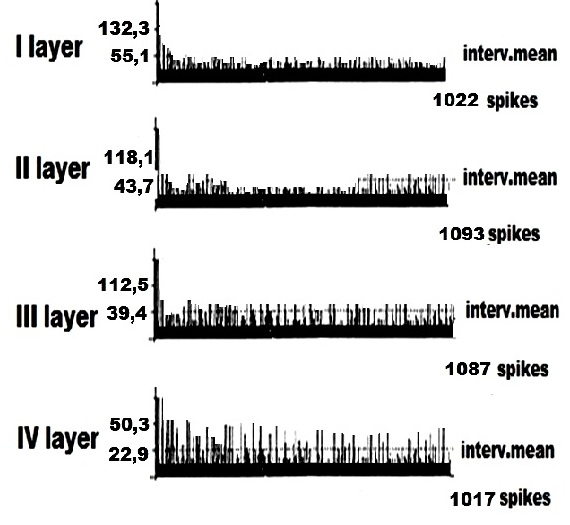
Рисунок 2. Пример программного анализа внеклеточной фоновой электрической активности одиночных пирамидных нейронов I – IV cлоёв коры больших полушарий головного мозга у крыс с левосторонней латеральной гемисекцией спинного мозга. Представлена частота импульсов в реальном времени и средняя величина межимпульсных интервалов.
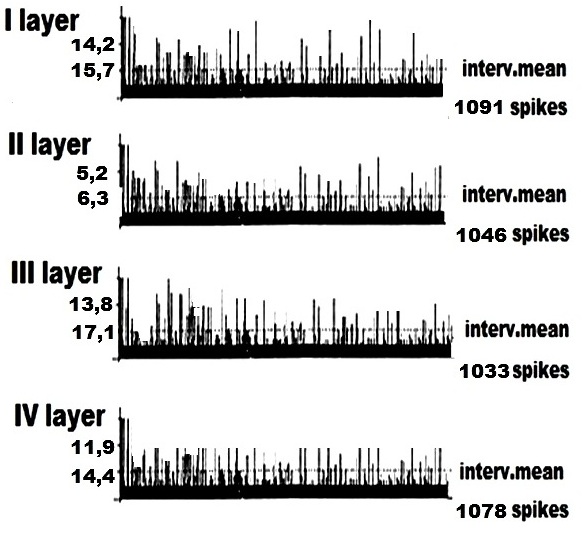
Рисунок 3. Пример программного анализа внеклеточной фоновой электрической активности одиночных пирамидных нейронов I – IV cлоёв коры больших полушарий головного мозга у крыс получавших систематические инъекции гормона щитовидной железы – тироксина. Представлена частота импульсов в реальном времени и средняя величина межимпульсных интервалов.

Рисунок 4. Пример программного анализа внеклеточной фоновой электрической активности одиночных пирамидных нейронов I – IV cлоёв коры больших полушарий головного мозга у крыс с левосторонней латеральной гемисекцией спинного мозга, получавших систематические инъекции гормона щитовидной железы – тироксина. Представлена частота импульсов в реальном времени и средняя величина межимпульсных интервалов.