

Введение
Проблема состояния восстановительных процессов при повреждениях спинного мозга (CМ) у млекопитающих при воздействии препаратов различного происхождения является одним из актуальнейших вопросов современной биологии и медицины [1 – 3]. Однако стойкость соматических и вегетативных нейрогенных нарушений является причиной инвалидизации большинства больных с поражением СМ, во время которых нарушается проведение нервных импульсов [4 – 7]. Наши исследования были направлены на поиск оптимального средства, стимулирующего и благоприятствующего росту волокон повреждённых путей СМ [8 – 10]. Известно, что вода покрывает три четверти поверхности Земли. Она составляет значительную часть живого организма (60 – 70 % тела человека, 90 % – растений). Форма её нахождения обуславливается соотношением давления и температуры: например, туман, облака, дождь, иней, град, снег [11]. Также известно, что вода является преобладаю¬щим компонентом содердания живой клетки (85 %), средний молекулярный вес равен 18; число молекул воды на единицу ДНК составляет 1, 2 * 10-7. Вода служит естественным растворителем для минеральных ионов и других веществ, а также дисперсионной средой, играющей важнейшую роль в коллоидной системе протоплазмы [12]. Тело человека почти на 63 – 68 % состоит из воды. Почти все биохимические реакции в каждой живой клетке – это реакции в водных растворах. С водой удаляются из нашего тела ядовитые шлаки; вода, выделяемая потовыми железами и испаряющаяся с поверхности кожи, регулирует температуру нашего тела. Представители животного и растительного мира содержат такое же обилие воды в своих организмах. Меньше всего воды, лишь 5 – 7% веса, содержат некоторые мхи и лишайники. Большинство обитателей земного шара и растения состоят более чем на половину из воды. Например, млекопитающие содержат 60 – 68 %; ры-бы – 70 %; водоросли – 90 – 98 % воды [13]. Имеется ещё один вид воды, отличающийся по физическим свойствам от обычной воды, – это омагниченная вода (ОМВ). Такую воду получают с помощью магнитов, вмонтированных в трубопровод, по которому течет вода. ОМВ изменяет свои физико – химические свойства: скорость химических реакций в ней увеличивается, ускоряется кристаллизация растворённых веществ, увеличивается слипание твёрдых частиц примесей и выпадение их в осадок с образованием крупных хлопьев (коагуляция). Омагничивание успешно применяется на водопроводных станциях при большой мутности забираемой воды. Она позволяет также быстро осаждать загрязненные промышленные стоки. С биологической точки зрения ОМВ является биогенным фак¬тором, стимулирующим мочеотделение, снижение артериального давления, изменение фармакологического действия ряда лекарств и других закономерностей протекания процессов в живом организме [14 – 16].
Особенностями структурированной (талой) воды (СВ) является то, что она свободно диффундирует через клеточную мембрану, способствуя быстрому проникновению внутрь клетки растворённых в ней веществ [17].
Йодированная вода (ИВ) обладает широким спектром действия на са¬мые разнообразные процессы в живом организме, что отражено в ряде научных исследований [18 – 20].
В данных сериях экспериментальных исследований было проведено изучение воздействия ОМВ, СВ и ИВ на изменение внеклеточной вызванной электрической активности (ВА) одиночных мотонейронов (МН) СМ крыс при экспериментальном спинномозговом повреждении средней степени тяжести – левосторонней латеральной гемисекции (ЛЛГМС). Данные биогенные факторы (БФ) использовались нами в целях стимуляции обменных процессов в повреждённых клетках, со стимуляцией роста аксонов, по которым восстанавливалась проводимость импульсов от периферии к коре головного мозга.
Материал и методика
Эксперименты поставлены на 50 белых крысах – самцах, массой (210 – 230 г), разделённых на следующие подопытные группы: первая – 10 экземпляров – интактные животные; вторая – 10 экземпляров – животные с ЛЛГМС СМ на уровне Т8 – Т9; третья – 10 экземпляров – животные с ЛЛГМС СМ на уровне Т8 – Т9 получавшие в течение 1 месяца ежедневно ОМВ (дозировка – 100 мг/кг массы животного, каждое животное индивидуально); четвёртая – 10 экземпляров – животные с ЛЛГМС СМ на уровне Т8 – Т9 получавшие в течение 1 месяца ежедневно СВ (дозировка – 100 мг/кг массы животного, каждое животное индивидуально); пятая – животные с ЛЛГМС СМ на уровне Т8 – Т9 получавшие в течение 1 месяца ежедневно ИВ (дозировка – 100 мг/кг массы животного, каждое животное индивидуально). Омагничивание воды проводили посредством специальной установки «Tauras – 100i»; СВ получали каждодневным замораживанием водопроводной воды в холодильной установке марки «Okai», Япония; ИВ получали посредством добавления в водопроводную воду калия йодида из расчёта: 5 капель KI на 250 мл воды. После проведения клинических наблюдений и дачи препаратов на всех 5 группах животных были поставлены электрофизиологические эксперименты. Производили экстраклеточную регистрацию ВА одиночных МН вентрального рога СМ крыс при одиночном, надпороговом раздражении седадищного нерва нижней конечности. Отведение активности исследуемых МН проводили стеклянным микроэлектродом с диаметром кончика 1 – 2 микрон, заполненных 2М раствором NaCl, в дорзо – вентральном направлении в сером веществе передних рогов СМ в области МН (IX пластина по Рекседу). Регистрацию ФА МН СМ проводили с помощью спе-циально разработанной программы, обеспечивающей в режиме on-line селекцию спайков посредством амплитудной дискриминации спайка для выбора необходимого режима записи ВА одиночного МН. Анализ полученных данных осуществляли по алгоритму, подробно описанному в наших предыдущих статьях [8]. Более подробно с программной методикой наших экспериментов можно ознакомиться в наших работах [21, 22].
Результаты и обсуждение
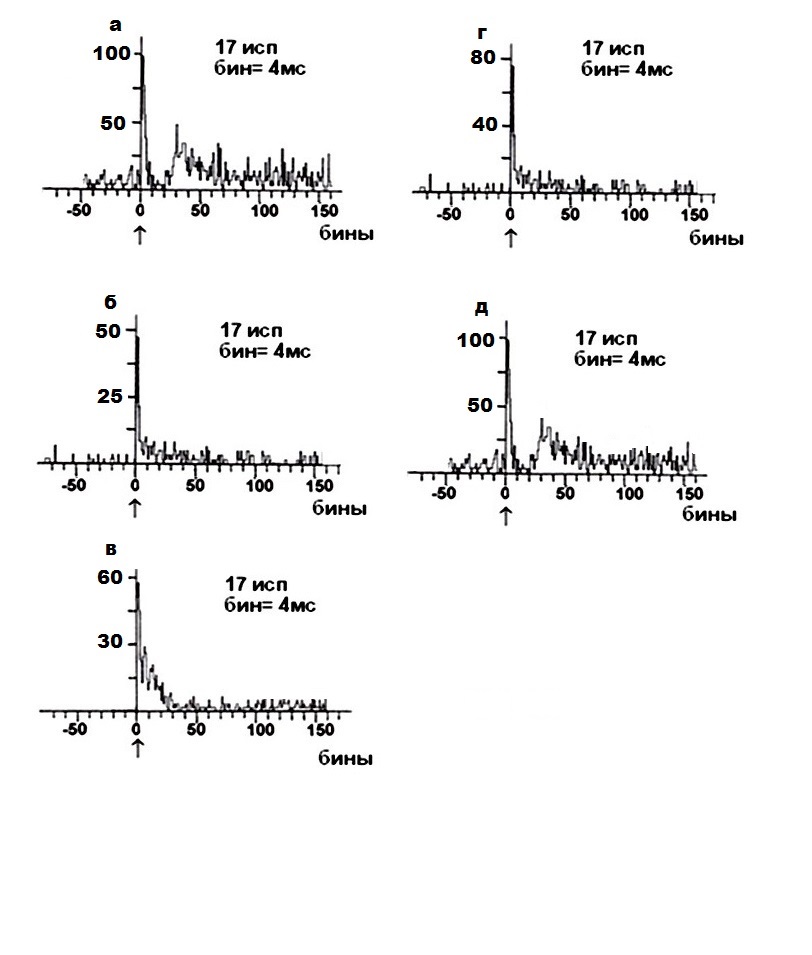
Рис. 1. Суммированные постстимульные гистограммы внеклеточной вызванной активности одиночного мотонейрона (глубина 1200 мкм) вентрального рога спинного мозга крыс в норме (а); одиночного мотонейрона (глубина 1200 мкм) вентрального рога спинного мозга крыс при левосторонней латеральной гемисекции спинного мозга (б); одиночного мотонейрона (глубина 1200 мкм) вентрального рога спинного мозга у крыс с левосторонней латеральной гемисекцией спинного мозга, получавших в течение 1 месяца омагниченную воду (в); одиночного мотонейрона (глубина 1200 мкм) вентрального рога спинного мозга у крыс с левосторонней латеральной гемисекцией спинного мозга, получавших в течение 1 месяца структурированную воду (г); одиночного мотонейрона (глубина 1200 мкм) вентрального рога спинного мозга у крыс с левосторонней латеральной гемисекцией спинного мозга, получавших в течение 1 месяца йодированную воду (д). Ордината – процент импульсов (в бинах) от числа проб, абсцисса – последовательность бинов.
На рис. 1 демонстрируется пример кумулятивной престимульной гистограммы внеклеточной ВА одиночного МН СМ крыс (глубина 1200 мкм) в норме (рис. 1, а); у животных с ЛЛГМС СМ (глубина 1200 мкм, рис. 1, б); у животных с ЛЛГМС СМ, получавших ОМВ в течение 1 месяца (глубина 1200 мкм, рис. 1, в); у животных с ЛЛГМС СМ, получавших СВ в течение 1 месяца (глубина 1200 мкм, рис. 1, г); у животных с ЛЛГМС СМ, получавших ИВ в течение 1 месяца (глубина 1200 мкм, рис. 1, д). Как видно из рис. 1, б, последствия спинномозгового повреждения (ЛЛГМС) проявляются в виде урежения регулярного разряда внеклеточной ВА одиночного МН СМ крыс и наблюдается явление трансформации регулярного разряда ВА одиночного МН в патологический «пачечный» тип разряда данного МН. При действии ОМВ (рис. 1, в) не наблюдается нормализации картины внеклеточной ВА одиночных МН СМ у крыс и в наличии проявляется вышеописанный тип пачечного разряда одиночных МН СМ. При действии СВ (рис. 1, г) наблюдается явление частичной трансформации пачечного типа ФА одиночного МН СМ в регулярный тип разряда, но, впоследствии, преобладает пачечный тип разряда МН. При действии ИВ (рис. 1, д) наблюдается резкое учащение постстимульного ответа МН, характеризующееся учащением постстимульного вызванного импульсного потока, с явной трансформацией внеклеточной ВА в регулярный тип разряда, сопровождающееся исчезновением пачечной активности.
Анализируя проведенные исследования можно прийти к выводу о том, что в целом имеется положительный эффект от применения ИВ при органических повреждениях СМ крыс средней степени тяжести типа ЛЛГМС и наблюдается наличие стойких положительных результатов. Проведенные экспериментальные исследования позволяют считать, что после ЛЛГМС СМ, проведенной на уровне Т8 – Т9, наблюдается постепенная картина нормализации нарушений опорно – локомоторных функций и нагляднее всего это проявляется у крыс 3 – ей группы (в течение 16 – 19 дней). Таким образом, полученные результаты данного исследования свидетельствуют об эффективном действии ИВ на изменение внеклеточной ВА одиночных МН СМ при ЛЛГМС.
Ранее нами было описано действие СВ, ИВ и ОМВ в изменении внеклеточной фоновой электрической активности одиночных МН СМ крыс при экспериментальном гипотиреозе [23], а также микроэлектрофизиологическое исследование влияния малых доз комплекса минеральных вод России на изменение внеклеточной фоновой электрической активности одиночных МН СМ крыс в условиях экспериментального гипертиреоза [24]. На данном этапе отсутствуют данные относительно применения различных БФ при спинальных повреждениях.
Учитывая результаты ранее проведённых исследований по изучению действия СВ и ИВ [25 – 29], а также данные проведенного нами исследования, высказывается предположение о протекторном действии ИВ на внеклеточную ВА одиночных МН СМ при ЛЛГМС.
Литература
1. Андреасян А. С., Хачатрян Т. С. Влияние лидазы на проводимость повреждённого спинного мозга. Ж. Вестник МАНЭБ, 2003, т. 8, № 7, с. 206 – 210.
2. Марченко З. И., Матинян Л. А., Бабаханян М. А., Хачатрян Т. С., Киприян Т. К. Роль солодки (Glychyrrhiza Glabra L.) в изменении фоновой электрической активности пов¬реж¬дённых травмой седалищного нерва интернейронов и мотонейронов спинного мозга крыс. Ж. Вестник МАНЭБ, 2003, т. 8, № 4, с. 150 – 153.
3. Киприян Т. К., Чавушян В. А., Матинян Л. А., Хачатрян Т. С. Влияние дексаметазона на электрическую активность нейронов в повреждённой рефлекторной дуге спинного мозга. Ж. Медицинская наука Армении, 1998, т. 38, № 3 – 4, с. 23 – 29.
4. Киприян Т. К., Хачатрян Т. С., Матинян Л. А., Чавушян В. А. Электрическая активность нейронов спинного мозга крыс при действии тироксина, гипертиреозе и гипотирео¬зе. Ж. Вопросы теоретической и клинической медицины, 1999, т. 2, № 7, с. 50 – 54.
5. Galoyan A. A., Kipriyan T. K., Sarkissian J. S., Matinyan L. A., Chavushian V. A., Andreasyan A. S., Khachatryan T. S. Changes of the injured rat’s spinal cord neurons electrical activity under action of dexamethasone and hypothalamic neuropeptide hormone. In: 38th Annual scientific meetigs of International Society of Paraplegia, Copenhagen, Denmark, 1999, p. 85.
6. Galoyan A. A., Kipriyan T. K., Sarkissian J. S., Sarkissian E. J., Grigorian Yu. Kh., Sulkhanyan R. M., Khachatryan T. S. Protection against neuronal injury by hypothalamic peptides and dexamethasone. J. Neurochemical Research, 2000, v. 25, № 12, USA, рр. 1567 – 1578.
7. Agasian A. L., Amiryan S. V., Kipriyan T. K., Sarkissian J. S., Sarkissian E. J., Sulkhanyan R. M., Avetisyan Z. A., Khachatryan T. S. Isolated and conjoint with hormones action of Vippera raddei venom on spinal cord single neuron activity in norm and pathology. In: 4th Asian herpetological conference, 2000, Chengdu, China, July 16 – 20, p.1.
8. Хачатрян Т. С., Матинян Л. А., Андреасян А. С., Киприян Т. К. Роль тироксина в изменении электрической активности интернейронов и мотонейронов повреждённого спинного мозга крыс. Ж. Вопросы теоретической и клинической медицины, 2002, № 1, с . 40 – 45.
9. Киприян Т. К., Амирян С. В., Аветисян З. А., Хачатрян Т. С. Протекция дексаметазоном острой нейродегенерации интернейронов спинного мозга, вызванной змеиными ядами. Ж. Вестник МАНЭБ, 2002, т. 7, № 4, с. 155 – 158.
10. Киприян Т. К., Амирян С. В., Андреасян А. С., Хачатрян Т. С. Эффекты ком¬би¬ни¬ро-ван¬ного воздействия малых доз змеиных ядов и гормонов на электрическую активность интернейронов спинного мозга крыс. Ж. Вестник МАНЭБ, 2002,т. 7, № 6, с. 157 – 159.
11. Гаврюченков Ф. Г., Курочкин М. И., Потехин А. А., Рабинович В. А. Химия. Справочное руководство. Изд. «Химия», Л., 1975, с. 20.
12. Де Робертис Э., Новинская В., Саэс Ф. Биология клетки. Изд. «Мир», 1967, с. 35 – 37.
13. Спенглер О. А. Слово о воде. Л., Изд. «Гидрометеоиздат», 1980, с. 115.
14. Мамаев В. А., Мухин Ю. В., Кромм Д. С. Врачеватели древности: экскурс в историю. М., 1988, Изд. «СПС», с. 12 – 17.
15. Larsson E. M. Movement of water molecules and blood flow show the details of brain. J. Lakartidningen, 2008, № 105, v. 47, pp. 3430 – 3436.
16. Jacob M., Sutton B. P. Algebraic decomposition of fat and water in MRI. J. IEEE Trans. Med. Imaging., 2009, № 28, v. 2, pp. 173 – 184.
17. Lombardo T. G., Giovambattista N., Debenedetti P. G. Structural and mechanical pro¬per-ti¬es of glassy water in nanoscale confinement. J. Faraday Discuss., 2009, № 141, pp. 359 – 376.
18. Li S. M., Zhang G. H., Sun F., Wang P. H., Zhang Z. Z., Li X. W., Li S. H. Field study on the change of urinary iodine levels among family members with iodine content of 5 – 150 microg/L in drinking water before and after non-iodized salt intervention. J. Zhong-hua Liu Xing Bing Xue Za Zhi (China), 2008, № 29, v. 8, pp. 767 – 770.
19. Ren Q., Fan J., Zhang Z., Zheng X., Delong G. R. An environmental approach to cor¬rec-ting iodine deficiency: supplementing iodine in soil by iodination of irrigation water in remote areas. J. Trace Elem. Med. Biol., 2008, № 22, v. 1, pp. 1 – 8.
20. Weng H. X., Yan A. L., Hong C. L., Qin Y. C., Pan L., Xie L. L. Biogeochemical transfer and dynamics of iodine in a soil-plant system. J. Environ. Geochem. Health, 2008, № 18, p. 10 – 16.
21. Андреасян А. С., Матинян Л. А., Хачатрян Т. С. Роль тироксина в изменении фоновой электрической активности пирамидных нейронов коры больших полушарий головного мозга крыс при органическом повреждении спинного мозга. Ж. Вестник МАНЭБ, 2004, № 3, с. 142 – 145.
22. Киприян Т. К., Матинян Л. А., Андреасян А. С., Хачатрян Т. С. Влияние окситоцина и вазопрессина на электрическую активность нейронов спинного мозга крыс. Ж. Нейрохимия, 2005, т. 22, № 1, с. 57 – 65.
23. Хачатрян Т. С. Роль структурированной, йодированной и омагниченной воды в изменении фоновой электрической активности одиночных мотонейронов спинного мозга крыс при экспериментальном гипотиреозе. Ж. Мед. наука Армении, 2010, № 1, т. L, c. 56 – 64.
24. Хачатрян Т. С. Микроэлектрофизиологическое исследование влияния малых доз комплекса минеральных вод России на фоновую электрическую активность одиночных мотонейронов спинного мозга крыс в условиях экспериментального гипертиреоза. Ж. Мед. Наука Армении, 2009, № 3, т. XLIX, С. 75 – 84.
25. Guilherme M. R., Moia T. A., Reis A. V., Paulino A. T., Rubira A. F., Mattoso L. H., Muniz E. C., Tambourqi E. B. Synthesis and water absorption transport mechanism of a pH-sensitive polymer network structured on vinyl-functionalized pectin. J. Biomacromolecules, 2009, № 10, v. 1, pp. 190 – 196.
26. Curnow P., Booth T. G. The transition state for integral membrane protein folding. J. Proc. Natl. Acad. Sci. U. S. A., 2009, № 106, v. 3, pp. 773 – 778.
27. Bulliyya G., Dwibedi B., Mallick G., Sethy P. G., Kar S. K. Determination of iodine nutrition and community knowledge regarding iodine deficiency disorders in selected tribal blocks of Orissa, India. J. Pediatr. Endocrinol. Metab., 2008, № 21, v. 1, pp. 79 – 87.
28. Матинян Л. А., Нагапетян Х. О., Андреасян А. С., Киприян Т. К., Хачатрян Т. С. Об усилении некоторых целебных воздействий на человека и животных. Ж. Вестник МАНЭБ, 2007, т. 12, № 4 (вып. 2), с. 157 – 159.
29. Матинян Л. А., Нагапетян Х. О., Киприян Т. К., Авакян А. Э., Хачатрян Т. С. Особенности влияния структурированной воды на электрокардиограмму крыс в норме и при экспериментальном гипертиреозе. Ж. Вестник МАНЭБ, 2007, т. 12, № 4 (вып. 2), с. 184 – 186.