

УДК 581.1
М.М. Богдан
Институт физиологии растений и генетики Национальной академии наук Украины, Киев
Обзор посвящен анализу отдельных аспектов исследованию электрогенных Н+-насосов растительных клеток, роль калия и кальция в процессах транспорта ионов, калиевых- и кальциевых каналов, дальнего транспорта ионов в растениях и их локализации в органах и клеточных структурах. Роль микроэлементов в метаболизме растений.
Исследования электрогенных Н+-насосов растительных клеток.
Изучению механизма регуляции электрогенного транспорта посвящены многочисленные исследования [32, 33, 34, 40, 61, 66, 89, 120, 121, 134]. Окислительно-восстановительные (редокс) реакции в клетках живых организмов происходят, когда электрон (ē) переносится от донорных субстратов к акцепторным. Исследования редокс-реакций в течение последних десятилетий, сфокусировали свое внимание на митохондриях, хлоропластах и эндоплазматическом ретикулуме. Однако транспорт ē происходит также на плазматической мембране (ПМ) как в растительных, так и в животных клетках. Исследователи предполагали, что поглощение анионов и сопутствующая стимуляция дыхания контролируются цепью цитохромов на мембране. Исследования в направлении анион связанного дыхания пролили свет на локализацию и значение ПМ редокс-активности [121]. Многие из них взялись за развитие систем для анализа редокс-активностей на ПМ, очистки и определения компонентов, отвечающих за эту активность. Активный же транспорт Н+ в плазмалемме растительных клеток может осуществляться Н+-АТФазой или редокс-системами. Исследованиями было показано, что на роль электрогенного протонного насоса плазмалеммы кроме Н+-АТФазы может претендовать редокс-цепь плазмалеммы [32, 33, 34, 134], составляющими компонентами которой во фракциях плазматических мембран является флавины, цитохромы b и с и выделенный белковый комплекс, обладающий НАДН-оксидазной активностью. Также выявлены редокс-реакции в плазмалеммы, как в фотосинтезирующих, так и гетеротрофных клетках [33]. На счет участия редокс-цепи в работе электрогенного Н+-насоса плазмалеммы и формировании мембранного потенциала клеток, то единого мнения у исследователей нет. Результаты опытов, моделирующих работу редокс-цепи плазмалеммы растений in vitro, показали, что добавление НАДН в среду вызывает у мембранных везикул клеток колеоптиль кукурузы генерацию мембранных потенциалов («плюс» внутри везикул), по знаку и величине с АТФ-зависимым потенциалом на тех же везикулах [34].
Гипотеза о том, что электрогенный транспорт Н+ через плазмалемму клеток осуществляется с участием редокс-цепей впервые была высказана в 1978 году Новаком и Иванкиной [32]. Конечным акцептором электронов, которые переносятся редокс-цепью, при этом выступает, очевидно, О2. Например, на везикулах плазматических мембран корней хлопчатника генерация мембранных потенциалов («плюс» внутри везикул) происходит, когда донором электронов внутри везикул выступает аскорбат, а их акцептором внешне везикул – феррицианид. Редокс-цепь осуществляет электрогенный транспорт Н+ с растительной клетки [46], деполяризация плазмалеммы в ходе феррицианидредуктазной реакции дает основания считать, что феррицианид, конкурентно перехватывая электроны в редокс-цепи, препятствует выполнению данной системой функции электрогенного протонного насоса. Поскольку восстановление феррицианида происходит внеклеточно, оно должно приводить к нарушению трансмембранного разделения ē и Н+. Следовательно, выход Н+ из клетки сопровождается лишь закислением внешней среды. Исследователи предполагают, что восстановление феррицианида и подкисление среды, которое наблюдается при этом, обусловлено двумя механизмами: первичным транспортом только ē к феррицианиду, что приводит к деполяризации мембраны (за счет увеличения концентрации Н+ в цитоплазме), и последующим выходом Н+ осуществляемым Н+-АТФазой, которое активируется внутриклеточным подкислением и деполяризацией. Это подтверждают, в частности, результаты опытов с ингибитором Н+-АТФазной активности эритрозином В, который ингибирует то, что подкисление феррицианидом, влияет непосредственно на процесс восстановления феррицианида. Следует отметить, что сегодня особенно большое внимание уделяется исследованию роли редокс-системы в создании градиента электрохимических потенциалов Н+ ионов. Обобщая результаты исследований биоэлектрогенеза у высших растений, Опритов с соавторами [34] подчеркивают, что изначально при исследовании природы электрогенеза и транспорта ионов в растительных клетках считалось, что Н+-АТФаза является единственной электрогенной системой плазмалеммы. На сегодня же достоверно известно, что протон-движущая сила – градиент электрохимических потенциалов Н+ ионов образуется не только благодаря функционированию протонной Н+-АТФазы, но и редокс-системы, исходным звеном электрогенного транспорта ионов в которой есть восстановлены пиридиннуклеотиды НАДН2 и НАДФН2 и цитохромы b и с. Кроме того, предполагается существование целого ряда отдельных редокс-активностей с разной специфичностью к пиридиннуклеотидам. Авторы подчеркивают что, гипотеза о том, что электрогенный транспорт Н+ через плазмалемму клеток осуществляется с участием редокс-цепей [32]. В качестве теста на активность редокс-системы ими было предложено использование реакции восстановления феррицианида калия (ФГК) органами растений и установлено, что внеклеточное восстановление К3[Fe(CN)6] связано с глубокой деполяризацией плазмалеммы. Но в связи с отсутствием единого мнения и противоречащими высказываниями отдельных исследователей относительно участия редокс-цепей в работе электрогенного насоса и формировании метаболической составляющей электрохимического потенциала (Еm) Опритов с соавторами [34] обращают внимание на то, что первоочередной задачей при изучении электрогенеза растительных клеток является исследование соотношения между АТФ-и редокс-зависимым выделением Н+-ионов корнями растений при разных условиях выращивания.
Начало детального исследования способности растений к активности Н+-секреции и смещение рН питательного раствора относится к 20-40 годов прошлого века, когда в экспериментах стали использовать стеклянные рН-электроды и методы ионометрии [6]. В этот период S. Struger [12] высказал идею о влиянии ростовых веществ на кислотно-щелочные градиенты в тканях растений, на основе чего была разработана концепция так называемого «кислого роста». Суть ее заключается в том, что ауксины индуцируют процесс растяжения клеток путем активации транспорта протонов и их выхода из цитоплазмы в свободное пространство клеточных стенок. Снижение величины рН при этом способствует разрыхления и смягчению клеточных оболочек и растяжения клеток, т.е. растущая за экзогенной действия ауксина секреция протонов, которая происходит при участии АТФазы плазмалеммы, играет главную роль в росте клеток растяжением. Однако позже было установлено, что существование ауксин-зависимой активации Н+-помпы и роста клеток растяжением наблюдается не всегда и поэтому сейчас теория «кислого роста» ставится под сомнение. Обобщение данных литературы по этому вопросу приведены в работах [28, 137]. Их авторы отмечают, что отрицание противников теории «кислого роста» базируются на том, что кинетика секреции протонов часто не совпадает с кинетикой роста клеток растяжением. Кроме того, результатами многочисленных исследований показано существование аддитивного действия ауксина и органических кислот на выделение Н+ и рост растяжением. Поэтому исследование о зависимости процессов роста и выделения протонов клетками растений продолжается. Этому способствует появление новых методов исследования электрогенеза растений и изменение представлений о роли АТФазы в секреции протонов. На сегодня важны результаты, полученные при изучении роли ионных каналов в трансдукции ауксинового сигнала. Авторами этой работы [28] сделан вывод о том, что ИУК стимулирует выход протонов не прямо, а через последовательную активацию анионных, кальциевых и Са-зависимых Коut-каналов. Показано, что увеличение содержания Са2+ в цитоплазме активирует Са-зависимые К-оut каналы и выход К+ из клеток, что в свою очередь, способствует росту активности Н+-АТФазы и гиперполяризации плазмалеммы. Согласно Терещенко [45, 46], ИУК – индуцированный рост и выход Н+ из клеток при снижении рН следует рассматривать в связи с изменением интенсивности и направлением метаболических процессов. Так что вопрос о зависимости между электрогенным транспортом Н+ ионов и ростом клеток требует дальнейшего исследования.
Роль калия в электрогенезе растительных клеток.
Общеизвестно, что в растительных клетках калий находится в ионной форме и не входит в состав органических веществ, благодаря чему он характеризуется высокой подвижностью, легко реутилизуеться и играет важную роль в процессах транспорта ионов и водообмене растений. Наибольшее количество калия обнаружено в молодых тканях листьев, стеблей, почек, которые характеризуются высокой интенсивностью метаболических процессов. Особенно высокое содержание калия находится в тканях меристемы и камбия [38]. По данным [6], ионы калия относятся к минорным компонентам почвенного раствора или водной среды и, поэтому в наибольшем количестве накапливаются в растительных клетках по сравнению с другими ионами. Показано, что содержание обменного калия, который является основным источником питания растений, варьирует в почвах от 40 до 500 мг/кг, а 150 мг/кг является оптимальной его концентрацией [3]. Анализ литературных данных свидетельствует о том, что роли калия в жизнедеятельности растений были посвящены многочисленные физиолого-биохимические исследования. В 70-е годы прошлого века было известно, что калийные удобрения способствуют не только повышению урожайности сельскохозяйственных культур и качества растительной продукции, но и положительно влияют на устойчивость растений к различным грибковым заболеваниям, морозо- и устойчивости злаковых к полеганию. Кроме того, установлено, что при недостатке калия происходит угнетение процессов водообмена, фотосинтеза и синтеза белков, полисахаридов и жиров и активности целого ряда ферментных систем, в результате чего урожай растений снижается на 40 – 80 %. Однако, как справедливо отмечали А.С. Оканенко и Б.И. Берштейн [90], на основе полученных в 60-70 годы результатов физиолого-биохимических исследований нельзя было получить ответ на вопрос, принимает калий участие во всех перечисленных процессах, или он действует на один из них, нарушение которого и приводит к глубокому изменению всего обмена веществ в растениях. По Воробьеву [5], поворотным пунктом в исследованиях роли калия в растениях стало зарождение одного из фундаментальных направлений физиологии растений, а именно биоэлектрохимии растительной клетки, инициаторы которой предсказывали огромную роль биоэлектрических явлений в растениях и их непосредственная связь с поглощением и транспортом К+. Благодаря развитию биоэлектрохимии стало известно, что калий стимулирует активность белков-генераторов тока и то, что мембранным потенциалом растительных клеток управляют К+-каналы. Признание этого постулата стало возможным в 90-е годы прошлого века после объединения двух параллельно существующих направлений исследования ионного транспорта в растениях. Основой одного из них, который развивался со времен Д. Хогланда, X. Мондегорда, Д. Сабинина и обобщенный П. Митчелом и В. Скулачевым, было исследование преобразования химической энергии АТФ в электрохимический градиент протонов, который и используется на транспорт ионов. Во второй же, так называемой кинетической модели Эпстейна, энергетический преобразователь отсутствует, а поглощение и транспорт ионов рассматривается в связи с существованием двух механизмов, роль которых выполняют переносчики на плазмалеммы. Поскольку транспортный аппарат корневой системы растений в диапазоне механизма и адаптировался к низким концентрациям К+ (10-50 мкм), нитрата (10 мкм) и фосфора (3-5 мкм), и поскольку в ряду катионной стимуляции Н+-насосов калий занимает первое место (К+ > NH4+ > Rb+ > Na+ > Cl-), то считается, что одной из главных его функций в минеральном питании растений как одного из элементов триады NPК является то, что он необходим для стимуляции Н+-транспортного аппарата растительных клеток, генерирующих протон-движущую силу, которая расходуется на перенос нитратов, фосфатов, сахаров, аминокислот.
Итак, благодаря широкому использованию биоэлектрохимических методов исследования было показано [96], что транспорт К+ через плазмалемму осуществляется либо пассивно через К+-каналы по электрохимическому градиенту или вторично активно с потоками Н+ или Na+. Макроскопические же потоки К+ через поверхность клетки или протопласта находятся под контролем цитоплазматической активности Са2+, К+, Н+. Л.Н. Воробьев и Н.Н. Егорова [6], рассматривая концепцию активного поглощения ионов минерального питания, основанную на взаимосвязанном функционировании электрических Н+-насосов и потенциал зависимых К+-каналов, и физиологическую роль К+ как стимулятора Н+-насосов со стороны внешнего (питательный раствор) и внутреннего (цитоплазма клеток) среды, отмечают, что функционирование Н+-насосов находится под контролем не менее 3-4-х факторов: концентрации К+ в среде, накопление его в цитоплазме и вакуоли и дальнего транспорта К+ для обеспечения ростовых процессов. Стабильный транспорт К+ в сосуды ксилемы начинается при наличии 0,1 мМ К+ в питательном растворе, когда происходит существенная стимуляция Н+-насосов, хотя для интенсивных сортов оптимум К+ в среде может быть увеличен до 1-2 мМ, что означает переход от работы ферментативного механизма Н+, К+ насосов, в взаимосвязанное их функционирования с К+-каналами. Обобщающие литературные данные авторы [96] отмечают, что суть новой концепции о структурно-энергетической организацию мембран состоит в том, что мембранный потенциал создается особыми белками-генераторами тока, электрогенными Н+-АТФазами или отдельными звеньями редокс-системы, которые стимулируются калием, а также в том, что мембранным потенциалом управляют К+-каналы, которые являются составной частью белковых мембран и через которые происходит движение растворенных веществ в клетку и из клетки в окружающую среду. Важным свойством каналов является избирательность. Оно заключается в пропускании определенных ионов и способности к изменению конформационного состояния, определяющим в среде их активность. Выполнение К+-каналами своих функций обусловлено особенностями их строения, включая селективный фильтр и датчик напряжения и другие структуры, обеспечивающие селективность и активность их действия. Показано, что К+-каналы играют регуляторную роль не только в поглощении и выделении К+ из клеток, но и в тургорном движениях замыкающих клеток устьиц, в процессах растяжения и роста клеток, катионном питании клеток и внутриклеточном распределении К+ и генерации потенциала действия наряду с Са2+- и Cl- каналами.
При изучении роли калиевых каналов установлены некоторые различия между теми К+-каналами, находящихся в плазмалемме и тонопласте клеток корней, паренхиме ксилемы и замыкающих клеток устьиц листьев по главным параметрам, в частности таких как их количество, направленность движения К+ ионов в клетку (К+in-канал) или клетки (К+out-канал), время открытости и зависимость функциональной активности К+-каналов от величины мембранного потенциала на поверхности плазмалеммы и наличие К+ в среде, его кислотности и соотношения концентрации К+ с концентрацией других ионов. Полученные разными авторами данные свидетельствуют о значительных различиях между изучаемыми ими К+-каналами [72, 73, 74].
Поглощение и транспорт кальция в растительных организмах.
Долгое время, примерно до 60-х годов XX века поглощение кальция растениями рассматривалось как такое что, происходит по градиенту концентрации, т.е. благодаря пассивному транспорта. Еще Ю.Я. Мазель [27] в 1956 году, обобщая литературные данные, отмечал, что поглощение кальция различными растениями на протяжении вегетации имеет характер одновершинной кривой с максимумом в период цветения и плодоношения, что уже тогда можно было рассматривать как побочное свидетельства о существовании положительной зависимости между поглощением кальция и интенсивностью метаболических процессов. Теперь экспериментально доказана зависимость поглощения кальция от метаболизма. Кроме того общепризнанным является то, что трансмембранный перенос ионов может происходить пассивно по градиенту электрохимического потенциала иона и активно с затратой метаболической энергии, против градиента [4]. Активный транспорт ионов Са2+ и К+ может осуществляться с помощью редокс-систем плазмалеммы и ионных насосов АТФ-азной природы, роль которых сводится к созданию электрохимического потенциала ионов водорода на плазмалемме, по которому и другие ионы вторично попадают в клетки. По данным [29], в плазмалемме, тонопласте и аппарате Гольджи локализованы две системы поглощения Са2+. Одной из них является Са2+-АТФаза, которая характеризуется высоким сродством к ионам Са2+, но низкой способностью к его транспорта. При изучении эффективности поглощения катионов отрезанными корнями ячменя установлено, что поглощение Са2+ и Na+ в отличие от поглощения К+ является менее энергозависимой.
При рассмотрении данных о кальциевых каналах [133] отмечается, что кальций как основной элемент питания растений необходим для формирования структур, поддержки осмоса и осуществления сигнальных функций в клетках. Поступления Са2+ в клетки корня по градиенту концентрации обусловлено действием ионных каналов, и скорость поглощения его поддерживается на уровне 40 нМ/час г сырой массы корней. Скорость поглощения кальция бобовыми и разнотравьем значительно выше, чем зерновыми культурами. Чистое поглощения ионов Са2+ наиболее интенсивное в апикальной зоне (до 5 мм от кончика корня), а транспорт в ксилеме осуществляется через клетки эндодермы в области поясков Каспари. Поглощение может происходить против электрохимического градиента с участием Са2+ транспортной АТФазы или путем Са2+/Н+-антипорта. Са2+-каналы плазмалеммы корня включают катионные каналы с высокой проводимостью и зависящие от напряжения каналы с низкой проводимостью, где приток Са2+ ингибируется ионами La3+, Cd3+. Автор отмечает также существование неспецифических Са2+ проницаемых каналов, сформированных белками с фенилалкиламиннимы связями и имеют низкую общую проводимость [43].
По данным [5], в тонопласте высших растений существует 2 класса Са2+-каналов. Один из них регулируется инозит-1, 4,5 тиофосфатом, а второй электрическим напряжением. Открытие последнего индуцируется смещением электрохимического потенциала в положительную сторону. Активность же функционирования его не зависит от добавок инозит-1, 4,5 тиофосфата и варьирование цитоплазматического свободного Са2+. Он примерно в 20 раз более выборочный для ионов Са2+, чем для ионов К+ [87]. Однако согласно [44], тонопласт вакуоли содержит 2 протонные фосфогидролазы, различные переносчики, которые используют энергию протонного градиента или АТФ, а также несколько типов кальциевых, анионов и калиевых каналов. Показано также, что при низком уровне ионов Са2+ проводимость возрастает для тонопласта. Эта проводимость ионов К+ определяется работой быстрых вакуолярных каналов, в том числе анионных и потенциал зависимых Са2+-каналов. При увеличении концентрации свободного цитоплазматического кальция активность потенциал-зависимых Са2+-каналов уменьшается и мембранный потенциал смещается в сторону более отрицательных значений. При этом создаются условия для активации потенциал зависимых Са2+-каналов, которые могут ограничивать высвобождение вакуолярного Са2+. Важными являются данные, полученные при изучении активности Са2+-АТФазы и Са2+/Н+-антипорта в мембранах изолированных вакуолей фасоли. Они свидетельствуют о том, что при физиологической концентрации ионов Са2+ в растительных клетках в пределах 0,1 мкм, Са2+-АТФаза тонопласта является главной системой транспорта Са2+ [139]. Изучение транспорта ионов через тонопласт способствует выявлению механизмов осморегуляции, регуляции тургора и реализации других функций вакуоли и клетки в целом, но полученные разными авторами результаты кинетики ионных потоков достаточно противоречивыми. Возможно, это связано с тем, что цитоплазматические мембранные и вакуольные Са2+-каналы принимают участие во многих сигнальных системах, реакция которых на изменение условий окружающей среды носит для отдельных высших растений специфический характер. Как известно [86], изменение содержания свободного Са2+ в цитоплазме является основным звеном, которое связывает специфические типы трансдукции сигнала в растительной клетке. От активности Са2+-каналов зависят изменения уровня Са2+-индуцированной трансдукции сигнала в растительной клетке. Однако специфическая роль каждого из каналов плазмалеммы и тонопласта еще полностью не установлена. Кроме того, источником для передачи специфических сигналов является пулы ионов Са2+. Согласно Тарчевскому [43] у высших растений существует несколько типов Са2+-каналов, благодаря которым Са2+ движется через плазмалемму, тонопласт, мембраны эндоплазматического ретикулума, хлоропластов и ядер. Эти каналы имеют различные электрические характеристики и различную чувствительность к блокатора верапамила и ионов La3+. Сигнальную роль выполняют, главным образом, кальциевые каналы, которые активируются деполяризацией мембран от -140 мВ до менее отрицательных значений, очевидно обусловлено изменением конформации белков кальциевых каналов и их открыванием.
Поглощения и дальний транспорт ионов в растениях. Обменная адсорбция ионов.
Как известно, исследованию механизмов мембранного переноса ионов предшествовало изучение обменной адсорбции в клетках стенках. Наряду с этим параллельно достаточно активно проводились исследования дальнего транспорта ионов в растениях их локализации в органах и клеточных структурах. Отдельно большое внимание уделялось роли микроэлементов в растительном организме. Благодаря исследованиям обменной адсорбции в клеточных стенках было установлено, что поглощение ионов начинается с контактного обмена между ризодермой, клетки которая способна к образованию корневых волосков, и почвенным раствором и грунтовыми частями [5, 6]. В естественных условиях, когда растения получают необходимые минеральные вещества из почвы, их корни находятся в сложной системе биологических и физико-химических взаимоотношений между почвенными частицами, почвенным раствором и почвенными микроорганизмами и грибами. В этом случае ионы минеральных солей могут поступать в клетки корней как с почвенного раствора, так и в результате контактного обмена с почвенными частицами. Но поскольку большая часть питательных веществ находится не в почвенном растворе, а адсорбированная на грунтовых частицах, то в минеральном питании растений большую роль играют процессы обменной адсорбции, которая осуществляется благодаря тому, что клетки корней, как и почвенно-поглощающий комплекс ионов имеют свойства ионнообмеников. Ионообменные свойства клеток корней, обусловлены наличием в составе органах целлюлозы, гемицеллюлозы и пектиновых веществ, которые содержат отрицательно заряженные карбоксильные группы. В результате этого клеточные стенки концентрируют положительно заряженные катионы. Установлено, что поступление катионов и анионов в клеточные стенки ризодермы корней связано как с обменом Н+ на катионы внешней среды так с обменом анионов НСО-3 (ОН-) или анионов органических кислот на анионы минеральных веществ.
Согласно [31], контактный обмен ионов клеточной оболочки ризодермы осуществляется без перехода ионов в почвенный раствор. Тесный контакт между клеточными стенками ризодермы и почвенными частицами происходит благодаря отсутствию у ризодермы защитных покровных образований, в частности кутикулы, так и выделению волосками слизи. Обменная адсорбция ионов в клеточных стенках рассматривается, как первый самый быстрый этап поглощения элементов питания. Способность растений к обменной адсорбции и контактного ионообмена определяется обменной емкостью корней и зависит от химического состава корневых выделений и клеточных оболочек и поддерживается непрерывным синтезом новых веществ и их транспортом через цитоплазматическую мембрану. Итак, благодаря контакту оболочек соседних клеток существует единая система клеточных стенок, которая называется апопластом. По нему от клетки к клетке передвигаются различные вещества. Кроме того, в растениях функционирует единая система цитоплазмы клеток тканей и органов, которая называется симпластом. Ее функционирование обусловлено тем, что клеточные оболочки пронизаны порами диаметром до 1 мкм, через которые проходят тяжи-плазмодесмы, по которым также осуществляются межклеточные контакты. Центральную часть пор клеточных стенок занимают десмотрубочкы, вокруг каждой из них находится слой цитоплазмы, как и плазмодесмы соединен с цитоплазмой соседних клеток. Сами же десмотрубочкы, кроме того связанные с мембранами эндоплазматического ретикулума соседних клеток. Межмолекулярное пространство в клеточных стенках, где осуществляются процессы диффузии и ионной адсорбции включая поверхностью плазмалеммы, получил название водного свободного пространства (ВСП), которое занимает 5-10 % от объема растительных тканей и играет важную роль в поглощении и транспорте ионов [31].
Считается, что поступление ионов к свободному пространства апопласта происходит благодаря массовому потоку, диффузии и адсорбции. Главную роль в обогащении ионами СП играет процесс адсорбции, в основе которого лежит взаимное притяжение ионов и молекул клеточной стенки. Оно основывается на электростатических силах: отрицательно заряженные молекулы клеточной стенки (пектиновые вещества, глюкопротеиды) избирательно притягивают к себе катионы. В связи с этим содержание минеральных веществ в клетках не соответствует концентрации их во внешней среде. По этой причине, очевидно, в различных органах растений накапливается неодинаковое количество макро- и микроэлементов. Так, по данным [37], концентрация фосфора в тканях может превышать его содержание в питательном растворе почвы в 1-10 тысяч раз, калия и азота в 80 и 170 раз, а кальция и магния – в 56 и 7 раз соответственно. Это свидетельствует о наличии выборочного поглощения и накопления ионов в органах растений. Начало активного изучения этих процессов приходится на 50-60 годы прошлого века. В этот период под руководством П.А. Власюка благодаря использованию методов радиоактивных изотопов и эмиссионного спектрального анализа была установлена способность накапливать в органах отдельных микроэлементов в больших количествах при незначительном их содержании в почве. В плодах огурцов, например, обнаружено высокое содержание серебра, в томатах – циркония, в капусте – йода и молибдена, в чесноке – ванадия, а в зерне кукурузы – золота [10]. Выборочное поглощение и накопление ионов в клетках и органах свидетельствует о том, что минеральное питание растений в целом это сложная цепь биофизических, биохимических и физиологических процессов со своими обратными и прямыми связями, а также системами регуляции, многие из которых на сегодняшний день изучено недостаточно [31]. Но достоверным считается факт существования активного и пассивного транспорта веществ через плазмалемму [7, 14, 31]. Пассивный транспорт осуществляется путем диффузии по электрическому и концентрационному градиенту и происходит, тогда как концентрация их во внешней среде выше, чем в растениях. Активный транспорт осуществляется против электрохимического градиента с затратой метаболической энергии с участием электрогенных Н+-насосов (Н+ АТФазы, Na+ -, K+ -, Ca2 +-АТФазы и анионные АТФазы) и редокс-цепи, которые создадут электрический и химический (ΔрН) градиенты ионов Н+. Электрический потенциал ионов Н+ (мембранный потенциал) используется на транспорт катионов и анионов по электрохимическому градиенту против концентрационного. Обобщение литературных и экспериментальных данных относительно механизмов поглощения, транспорта и аккумуляции ионов в органах растений приведены в работах учащихся П.А. Власюка [31]. Авторы отмечают, что в поглощении, транспорте и метаболизме микроэлементов, как и макроэлементов, можно выделить следующие этапы
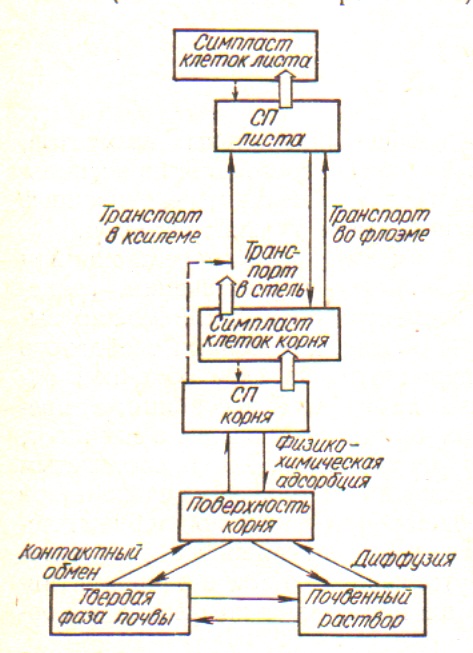 Рис. Общая схема поступления и транспорта ионов в растение
Рис. Общая схема поступления и транспорта ионов в растение
- Обогащение ионами СП (апопласта), которое происходит за счет обменной адсорбции, диффузии и пассивной физико-химической адсорбции;
- Активное преодоление ионами мембранного барьера и их поступлений в симпласт;
- Радиальное перемещение ионов по тканям корня и сосудистым проводящим пучкам;
- Вертикальное перемещение ионов по стеблям, черешкам и жилкам листьев;
- Включение наступающие извне ионов в метаболизм фотосинтезирующих клеток, их утилизация и реутилизации и отток в репродуктивные органы;
- Транспорт ионов и ассимилятов вниз по флоэме в корни (рис.).
Процесс выделения микроэлементов клетками корней во внешнюю среду является одним из важных вопросов, который требует детального исследования.
Важно отметить, что на сегодня получены данные о том, что выделение макроэлементов (азота, фосфора и калия) по их поглощения клетками корней зависит от факторов окружающей среды. Обнаружена суточная ритмичность поглощения и выделения клетками корней огурцов основных макроэлементов. Показано, что интенсивность поглощения и выделения во внешнюю среду азота, фосфора и калия зависит от их концентрации в питательном растворе [26]. Аналогичные данные были получены при изучении выделения бора во внешнюю среду при различной обеспеченности им растений риса. Показано, что интенсивное выделение бора клетками корней наблюдается при его высоком уровне в питательной смеси [144]. Но на сегодня недостаточно изученным является вопрос о радиальном транспорте ионов, а именно не известно каким образом ионы поступают в ксилему и как осуществляется ее загрузка [37]. Хотя хорошо известно, что путем диффузии и обменной адсорбции ионы поступают в клеточные стенки ризодермы и после, через кору паренхимы по апопласту и симпласту, до проводящих пучков. Движение ионов по апопласту путем диффузии и обменных процессов, ускоряется потоком воды, прекращается в эндодерме. Единственным дальнейшим путем транспорта веществ через эндодерму становится симпласт, где обеспечивается метаболический контроль их поступления. Наиболее активной мобилизации подвергаются вещества, содержащие азот, углерод, фосфор и в меньшей степени – серу, кальций, хлор. Другие ионы метаболического контроля практически не испытывают. Актуальны также исследования роли о симпластном транспорте веществ вакуолей, которые в определенной мере конкурируют с сосудами ксилемы за поглощение веществ. Предполагается, что регуляторная роль вакуолей зависит от степени насыщения их растворимыми веществами. Кроме того, вакуоли рассматриваются как запасной фонд питательных веществ при снижении их концентрации.
Открытие роли микроэлементов в метаболизме растений.
На сегодня известно, что в растительных и животных организмах находятся более 70 химических элементов. Одиннадцать из них макроэлементы (C, H, O2, N, S, P, Ca, Mg, K, Na) и микроэлемент (Se) составляют 99,95 % живого субстрата, и лишь 0,05 % его приходится на долю микроэлементов, которые играют чрезвычайно важную физиологическую роль [54].
Начиная с середины XIX века, среди физиологов существовало представление о том, что обнаруженные в органах растений в малых количествах (тысячные и десятитысячные доли процента) микроэлементы являются примесями, которые попадают в растительный организм благодаря наличию их в окружающей среде. Позже, когда с помощью физиологических методов исследований было установлено, что внесение в субстрат таких микроэлементов как бор, марганец, цинк, медь существенно повышает урожай сельскохозяйственных культур, их стали рассматривать как «стимулирующие» удобрения.
Первоначальные исследования по прикладных аспектах проблемы микроэлементов были направлены на определение оптимальных доз их внесения в почву при выращивании различных сельскохозяйственных культур.
Следует заметить, что исследование оптимальных доз внесения и соотношение микроэлементов в питательных растворах, которые интенсивно проводилось до середины ХХ века, продолжается и сейчас [42].
Анализ литературных данных свидетельствует о том, что в развитии учения о микроэлементах огромную роль сыграли два открытия. Первым из них, слишком важным для понимания физиологической роли микроэлементов было выявление того факта, что они входят в состав ферментов и, активируя их, играют чрезвычайно важную роль в обмене веществ. Поскольку почти все процессы синтеза и превращения веществ осуществляются с помощью ферментов, то считается, что микроэлементы являются основой основ жизни. На сегодня известно, что около 200 ферментов, то есть от 1/3 известного их количества активируется микроэлементами. Наиболее существенные успехи достигнуты при изучении стимулирующего действия таких микроэлементов, как железо, медь, марганец, молибден, магний. Обобщенные данные, приведенные в работе [54] свидетельствуют о том, что медь и железо являются компонентами таких ферментов, как бутирил-СоА-дегидрогеназа, ДПН-цитохром-с-редуктаза, дегидрогеназа фумаровой кислоты, сукциндегидрогеназа, нитритредуктаза (Escherichia coli), дегидрогеназа (Azotobacter vinelandiи), нитритредуктаза и гипонитритредуктаза (Neurospora crassa), нитритредуктаза и нитриоксидаза (Pseudomas stutzeri), молибден-нитратредуктазы (Escherichia coli, Neurospora crassa), марганец-гидроксиламинредуктазы и дегидрогеназы. Кроме того, марганец и магний активируют энзимы углеводного обмена (галактокиназу, фруктокиназы, глюкокиназы, гексозокиназу, фосфоглюкомутазы, киназу фосфоглицериновой кислоты, энолаза) и энзимы цинка лимонной кислоты (пировиноградную карбоксилазу и оксидазу, изолимонну и янтарную дегидрогеназу, а также разнообразные реакции фосфорилирования.
Другим не менее важным открытием в развитии учения о микроэлементах было выявление их способности образовывать с органическими соединениями органоминеральные комплексы (хелаты), которые играют важную роль в процессах внутриклеточного обмена. Показано, что в комплексных соединениях активность микроэлементов возрастает в сотни, тысячи и даже в миллионы раз по сравнению с их ионной формой. Происходит это благодаря образованию химических связей между ионами микроэлементов и отдельными группами молекул белков, или возникновению по соседству с ионом электростатических полей, сопровождается чаще всего изменением конфигураций белковых молекул и свойств ионов. Различные микроэлементы образуют хелаты неодинаковой устойчивости. Согласно [41] по убыванию величины образовывать хелаты металлов располагаются в следующем порядке: Cu > Ni > Co > Zn > Cd > Fe > Mn > Mg. Наиболее изученными являются соединения бора, меди, железа и молибдена с органическими веществами. Показано, что борная кислота легко образует хелаты по типу эфирной связи со спиртами, углеводами, оксикислотами и рядом других внутриклеточных соединений. Бор вступает даже в химическую связь с АТФ. Следует отметить, что открытие способности микроэлементов к образованию металлоорганических комплексов сыграло чрезвычайно важную роль не только для развития теоретических, но и для прикладных аспектов питания растений, а именно использование комплексных соединений для борьбы с хлорозом при недостатке железа и оптимизации питания растений при дефиците меди и других основных микроэлементов [36, 48]. На сегодня известно, что важным комплексным хелатообразуещим соединением является этилэндиаминтетрауксусная кислота (ЭДТА), которая образует комплексы почти со всеми металлами. Наиболее важными из них являются FeЭДТА, который используется для борьбы с хлороз при недостатке железа, и NaЭДТА, CuЭДТА, ZnЭДТА, MnЭДТА, MoЭДТА, которые применяются как микроудобрения для внекорневых подкормок. Важным хелатообразуещим, который используется в сельскохозяйственной практике для борьбы с хлорозом винограда и плодовых деревьев, также диэтилентриаминпентауксусная кислота (ДТПА). Установлено, что такие соединения как хлорофилл, гемины (составные части гемоглобина) и ферменты, которые содержат Fe, Cu, Mg, Co также есть хелатами. Как известно к железосодержащим ферментам относятся пероксидаза, каталаза и группа цитохромов, а те, которые содержат Cu, – полифенолоксидаза и аскорбиноксидаза [11]. Открытие того, что микроэлементы входят в состав огромного количества ферментов, как и их способности к комплексообразованию, способствовало отделению учения о микроэлементах в одну из важнейших проблем биологии. Согласно Уолтону [54], эти открытия вызвали революцию в аналитической химии и ознаменовали собой развитие новой области химии.
Локализация микроэлементов в клеточных структурах и их влияние на активность физиологических процессов.
Анализ литературных данных свидетельствует о том, что локализация микроэлементов в клеточных структурах и их влияния на активность физиологических процессов сыграл чрезвычайно важную роль для оптимизации системы питания различных сельскохозяйственных культур и совершенствования форм сложных комплексных удобрений. Среди основных достижений по этой проблеме следует назвать работы, которые выполнялись под руководством П.А. Власюка [8], Л.К. Островской [36], А.П. Кибаленко [21], З.М. Климовицкой [23, 24], Л.М. Кузьменко, М.Ф. Охрименко, В.И. Ивченко [18]. посвященные исследованию локализации микроэлементов Mn, B, Cu, Zn, Мо, Li в клеточных структурах и их влияния на активность физиологических процессов, и продуктивность сельскохозяйственных культур. Следует подчеркнуть, что наиболее значительные достижения при этом были установлены при изучении таких микроэлементов, как Mn, B, Cu, Zn.
Марганец. На сегодня известно, что марганец как железо, медь и молибден, имеет переменную валентность. Предвидя, что благодаря этому Mn играет важную роль в фотосинтетических реакциях, сначала изучали его локализацию в клеточных структурах листьев. По данным [9], основная масса марганца находится в цитозоле и хлоропластах, а при расчете на сухое вещество – в хлоропластах и митохондриях. Исследованиями [14] показано, что в гомогенатах тканей марганец находится преимущественно в ионном состоянии, а в живых тканях – в структурно связанной форме. В хлоропластах до 5 % его связано с белковой стромой, а остальные находятся в ионной или хелатной форме. Показано, что по содержанию Мn в разных сортах овощных культур наблюдаются существенные различия [20]. При изучении реакции растений на внесение марганцевых удобрений установлено [9], что дефицит марганца уменьшает прочность связи гран с ламелами стромы хлоропластов и способствует накоплению в них крахмальных зерен. Полное же его исключения из питательного раствора вызывает уменьшение гетерогенности белков хлоропластов и цитоплазмы и рост количества легкорастворимых белков. При дефиците марганца одновременно с угнетением интенсивности транспирации и ростовых процессов наблюдается уменьшение содержания нуклеиновых кислот в растительных клетках [24]. При этом происходит уменьшение содержания аскорбиновой кислоты в листьях и рост интенсивности выделения стрессового этилена [102]. В этих условиях меняется также гетерогенность основной генетической субстанции растений – хроматина. Показано, что через его дефицит уменьшается содержание общего белка и гистоновых и негистонових белков, а также ДНК и РНК в конденсированном и диффузном хроматине [12]. При избытке этого элемента в питательной смеси наблюдается нарушение двохспиральной структуры ДНК и уменьшение ее оптической плотности. Обнаружено [1, 97], что избыток марганца является одним из стрессовых факторов для растений, которые выращивались на кислых почвах. Увеличение его содержания в питательном растворе от 0,1 до 1 мМ вызывает уменьшение содержания в листьях фотосинтетических пигментов [91] и интенсивности фотосинтеза в листьях чувствительных к его содержимому сортов растений [97].
По данным [78], марганец выполняет определенную контролирующую функцию в клетках. С помощью метода ядерного-магнитного резонанса обнаружена способность растений поддерживать низкие концентрации марганца в цитоплазме клеток корней и неравномерное распределение его содержимого, как и кальция между цитоплазмой и вакуолями.
Изучение влияния марганца на продуктивность сельскохозяйственных культур показало, что положительная его действие в определенных концентрациях обусловила рост интенсивности фотосинтеза [9]. Исследованиями [49] показано, что внесение марганца в полевом севообороте на оподзоленном черноземе способствовало повышению урожая зерна озимой пшеницы на 11 %, а содержание белка и клейковины в нем на 1,4 % и 2,7 % соответственно.
Бор. Анализ литературных данных свидетельствует о том, что в 60-70 годах прошлого века довольно интенсивно проводились исследования влияния бора на фотосинтетическую активность растений. Экспериментами [88, 128, 131] показана положительная его действие на СО2-газообмен, содержание хлорофилла а, степень открытия устьиц, транспирации и аккумуляцию в листьях азота, пролина, сахаров и крахмала. Показано, что бор способствует увеличению количества и размеров хлоропластов. Большое значение для познания физиологической роли бора имеют исследования, выполненные под руководством А.П. Кибаленко [21]. В результате проведенных исследований установлено неравномерное распределение бора в корнях и надземных органах сахарной свеклы. Показано, что в листьях его содержание всегда выше, чем в корнях. В разных частях листьев количество бора также не одинакова. В паренхиме верхней части листа концентрация выше, чем в паренхиме нижней части и черешках и сосудисто-волокнистых пучках. Методом дифференциального центрифугирования наибольшее количество его обнаружено в хлоропластах, меньше в митохондриях, и совсем мало – в надосадочной жидкости. При борном голодании наблюдается дезорганизация системы тилакалоидов хлоропластов, увеличение в них количества пластоглобул. Его избыток также вызывает деструкцию межгранных тилаколоидов и размеров пластоглобул. Недостаток и избыток бора в питательной смеси приводит к нарушению анатомического строения листьев. При его дефиците наблюдается увеличение размера клеток нижнего эпидермиса и уменьшение размера и степени открытости устьиц, а при избытке – гипертрофия клеток эпидермиса и устьиц. Дефицит бора приводит к угнетению поглощения клетками корней глюкозы [100], и уменьшение содержания Са2+, связанного с плазматической мембраной. При этом наблюдалось увеличение количества свободного калия и кальция в апопласте [104].
Согласно [76, 94, 129], наиболее важными функциями бора в растениях является участие в транспорте сахаров, синтезе клеточных стенок и их лигинфикации, углеводном и РНК обмене, метаболизме ИУК и фенолов, процессах дыхания, формирования и поддержке функций клеточных мембран, тканей и органов растений.
Литературные данные свидетельствуют о том, что существенное влияние бора на продуктивность растений следует рассматривать в связи с его действием на активность ферментных систем. Так, выращивание растений табака на питательном растворе без бора показало, что подавление нарастание массы побегов и корней табака при снижении содержания в них бора, магния, кальция и калия и азота в 6-13 раз, сопровождалось снижением активности нитратредуктазы при росте в листьях содержания сахаров [69]. По данным [47, 130], бор, как и молибден положительно влияет на активность нитратредуктазы в листьях сахарной свеклы, масличного рапса, а также сои [95]. Внекорневые же подкормки растений способствует росту содержания бора, калия, магния, цинка, железа и меди наряду с уменьшением содержания марганца и аммония в листьях сои [118]. В опытах [111] с растениями подсолнечника, которые выращивались до 30-суточного возраста на смеси Хогленда-Арнона с различным содержанием бора, выявлено, что подавление активности пероксидазы происходило при наличии в питательной смеси содержания бора в концентрации 0,1 М и ее рост при повышении до 0,5 М.
В работе [122] приведены данные о влиянии обработки растений табака раствором борной кислоты в концентрации 0,5, 2,5, 10, и 25 мкм на активность ферментов фенилаланин аммиак-лиазы, полифенолоксидазы и пероксидазы, содержание бора и фенольных соединений. Показано, что бор в листьях находится преимущественно в 2-х формах: в комплексе с пектинами или фенольными соединениями в свободном состоянии. Действие бора зависило от примененной концентрации. Накопление фенольных соединений происходило по концентрации 0,5 и 25 мкм, а их окисление с 2,5 и 10 мкм. Показано, что в клетках растений бор находится в водорастворимой и нерастворимой форме [122]. Водорастворимый как нерастворимый бор, который связан с 2 молекулами боратов, локализованный не в протоплазме, а в клеточных стенках [103]. Значительное количество бора обнаружена в ксилеме, что указывает на его связь с транспирационным током. Ограниченным является содержание бора в флоэме [129].
По данным [63], большая часть бора (до 90 %) находится в клеточных стенках в составе боратов эфиров с ОН-группами, углеводов или глюкопротеидов. Он принимает участие в образовании полимеров с рибозой, маннозой, фруктозой. Содержание бора в клетках тесно коррелирует с пектином. Борсодержащий пектиновый полисахаридный комплекс ß-рамингалактурон является экскмозивним полисахаридом, находящимся в клеточных стенках. Включение глюкопротеидива и богатых пролином белков в сетчатую структуру клеточных стенок осуществляется только при наличии бора. В мембранах клеток бор стимулирует активность АТФазы и НАДН-оксидазы, усиливает гиперполяризацию мембран [104]. Предполагается, что бор, усиливая поглощение калия, активирует Н+-АТФазу, что и приводит к гиперполяризации мембран клеток корней [126]. Сейчас достаточно интенсивно проводятся исследования механизмов транспорта бора через плазматическую мембрану клеток корней. Согласно [144], поглощения бора растениями происходит пассивно при достаточном его обеспечении и активно при его низком содержании в питательной смеси. По борному дефициту наблюдается светлая окраска или хлороз молодых листьев риса и побурения их кончиков. При высоких концентрациях бора в питательной смеси происходит выделение его корнями растений. Однако у растений Sesamum indicum симптомы борного дефицита проявлялись и при высокой его концентрации, в частности, 21 мкг/г [117].
Важными являются данные [141] о том, что дефицит бора вызывает нарушение мембранных структур и тем самым приводит к росту интенсивности утечки калия из клеток и угнетение выделения протонов. В работе [62] отмечено снижение мембранного потенциала Еm после удаления бора из питательного раствора и его рост при добавлении бора. Предполагается [80], что бор может контролировать связывания ауксина с мембраной или влиять на сигнальную трансдукцию. Снижение содержания аскорбиновой кислоты в апопласте при росте в 4-10 раз железа в кончиках корней при дефиците бора свидетельствует о снижении восстановительного потенциала апопласта [64].
Предполагается, что участие бора как сильного комплексообразователя в обмене веществ следует рассматривать в связи с его влиянием на кальциевый метаболизм [30, 77].
Установлено [75], что в растениях и почве бор находится в свободном, полусвободном и связанном состоянии с соединениями, содержащими гидроксильную группу, а именно пентозы, гексозы, органическими кислотами (яблочная, лимонная, винная) и фенольными кислотами (галловая, протокатеховая, кофеиная).
Согласно [85], корни растений могут поглощать как свободную, так и связанную форму борной кислоты. При увеличении дозы борной кислоты в питательном растворе концентрация ее в листьях резко возрастает, но соотношение свободной и связанной формы остается постоянным. В корнях и стеблях большая часть борной кислоты находится в свободной форме. Максимальный рост растений пшеницы наблюдался при концентрации бора в питательной смеси равной 0,6 мкм [58]. Под влиянием повышенного поступления бора снижается скорость поглощения и накопления основных микроэлементов [106].
Вместе с тем выявлено, что при дефиците бора в питательной смеси в клеточных стенках возрастает содержание связанного бора при резком снижении его свободной формы [84].
В связи с тем, что большинство почв Украины характеризуются низким содержанием бора, важны исследования влияния способов внесения необходимых доз этого микроэлемента на формирование продуктивности сельскохозяйственных культур. Анализ литературных данных свидетельствует о противоречии результатов исследования разных авторов по решению этой проблемы. Так, согласно [52, 116], предпосевная обработка семян микроудобрениями является основным способом их применения. В частности, для выращивания высоких урожаев сахарной свеклы на легкосуглинистых почвах с содержанием бора 0,12-0,23 мг/кг рекомендуется вносить бор в почву. Из трех доз (1, 2 и 4 кг/га) самый высокий урожай был получен при норме внесения N160 P100 K300 + B4 [65].
Медь. Этот элемент занимает особое место в жизнедеятельности растений. В частности он входит в состав важнейших окислительных ферментов – полифенолоксидазы, аскорбиноксидазы, лактазы, дегидрогеназы, цитохромоксидазы, супероксиддисмутазы и других. Все ферменты, в состав которых входит медь, осуществляют реакции окисления переносом электронов с субстрата к молекулярному кислороду, который является их акцептором. В связи с этим валентность меди в окислительно-восстановительных реакциях меняется с двухвалентного до одновалентного состояния и обратно [2, 15].
Различные сельскохозяйственные культуры отличаются по чувствительности к дефициту меди. Растения, которые сильно реагируют на дефицит меди можно разместить следующим образом: пшеница > ячмень > овес > кукуруза > морковь > свекла > лук > шпинат > люцерна > белокочанная капуста. Средней чувствительностью к дефициту меди характеризуются картофель, помидоры, клевер красный, фасоль, соя.
Медь играет важную роль в процессах фотосинтеза. Она входит в состав белка пластоциану-донора электронов фотосистемы I, а также ферментов тирозиназы и лаказ, которые окисляют растительные фенолы. По данным [99], содержание меди в листьях значительно выше, чем в корнях. В листьях медь находится главным образом в хлоропластах [110]. За действием меди наблюдался рост активности пероксидазы, синтеза белков, углеводов и жиров [36], а также содержания хлорофилла в листьях риса [132] и дыни [19], реакции Хилла, интенсивности фотосинтеза и транспирации растений сафлора [112]. Показано [19], что предпосевное замачивание семян дыни в растворах меди, цинка и марганца способствовало повышению интенсивности дыхания проростков. Противоречивыми являются данные о влиянии различных доз меди на интенсивность некоторых других физиологических процессов. Так, при изучении влияния различных доз меди на флуоресценцию хлорофилла листьев смолянки и ярутки установлено [108] рост фотохимической активности хлоропластов при дозе меди 8 мкм и сильное ее угнетение при повышении дозы до 160 мкм. При исследовании же действия меди на выделение кислорода и флуоресценцию хлорофилла в опытах с хлоропластами гороха показано, что ингибирование исследуемых процессов происходит при повышении концентрации меди в питательном растворе в пределах от 5 до 20 мкм [110]. Однако при концентрации 160 мкм меди наблюдается ингибирование фотосинтеза и снижение содержания хлорофилла и кальция [109]. Избыток меди [113] влияет на равновесие между фотоингибированием и восстановлением, вызывая снижение равновесной концентрации активных центров фотосистемы II в освещенных листьях. При низкой же обеспеченности медью уменьшается вес всего растения и содержание в листьях хлорофилла, ослабляется активность ферментов полифенолоксидазы и повышается активность рибонуклеазы и пероксидазы. [71].
Исследования [105] влияния дефицита и избытка меди в питательном растворе на содержание сахаров и аскорбиновой кислоты в листьях и активность полифенолоксидазы и оксидазы, на сроки формирования зерна риса а также его урожайности и качество зерна, показало, что в молодых листьях предельное, достаточное и токсическое содержание меди составляет соответственно 3,9; 6,8; и 32 г/кг.
Согласно [56], избыток меди приводит к симптомам дефицита цинка в листьях, подавляет активность карбоангидразы, альдолазы и рибонуклеазы, цитохромоксидазы, полифенолоксидазы и нарастание массы кукурузы и подавляет образование дополнительных корней и ускоряет старение листьев риса [93]. Оптимум концентрации меди в среде для риса составил 0,065 мг/л [105]. При внесении 5, 10 и 20 кг/га меди, установлена положительная корреляция между урожаем зерна и содержанием меди в флагов листье. [123].
Влияние различных концентраций меди на содержание углеводов и белков в корневой вытяжке приведены также в работе [51]. Авторами показано, что обработка растений раствором меди (1 и 10 мг/л) приводит к увеличению резкого содержания белка в зерне.
Важны данные [59], которые свидетельствуют о связи между поглощением меди и изменением активности Cu-хелатредуктазы (АМХР), а также действия меди на восстановление хелатов железа. Выявлено, что добавление меди в питательный раствор активирует редокс-систему, плазматической мембраны клеток корней ячменя, облегчая тем самым восстановление хелатов меди и железа. При увеличении меди в питательном растворе от 0,05 до 0,1; 0,2; 0,4; и 0,8 мкм наблюдалось снижение концентрации цинка и азота в листьях огурцов [82]. Исследование распределения меди в органах растений показано, что высокая его концентрация находится в корнях и минимальная в зерновках [22].
Исследование влияния известкования и внесения CuO в дозе 0, 100, 300, 900 мг/кг почвы на урожай зерна кукурузы показало [57], что увеличение величины рН отрицательно влияло на поступление меди к корням растений и содержание ее в органах растений. Медь слабо влияет на поглощение К+ растениями при низкой его концентрации в питательном растворе, но подавляет этот процесс при высоких его концентрациях, блокируя транспортную систему на поверхности раздела симпласта и ксилемы [67]. Предполагается [101], что в аккумуляции, детоксикации и метаболизме меди, и кадмия, цинка, свинца и ртути задействованы фитохелатины – низкомолекулярные соединения с высоким химическим сродством к ряду металлов. Медные удобрения повышают сумму незаменимых аминокислот [25]. Обработка растений ячменя медью специфически влияет на активность пирофосфотазы протонной помпы [60]. Низкие концентрации меди увеличивают, а высокие уменьшают активность β-глюкозидазы. [55].
Цинк. Цинк, который является компонентом 800 ферментов, влияет на белковый, углеводный и фосфорный обмен растений. Он входит в состав таких ферментов как протеазы, аминопептидазы, карбоксипептидазы, дегидрогеназы, изомеразы, альдолазы, РНК и ДНК полимеразы, а в хлоропластах активирует карбоангидразы, катализирующей гидрирования диоксида углерода в бикарбонат:
Н2О + СО2 = Н+ + НСО3-
Исследованиями П.А. Власюка, А.К. Мицюры, Е.В. Рудаковой и других [50, 114] показано, что среди органов растений наибольшим содержанием цинка отличаются старые листья, стебли и семена, а среди клеточных структур – оболочки. Аналогичные данные получены [98] при изучении поглощения и аккумуляции цинка толерантными гиперакумулюючимы растениями. Однако в работе [115] отмечено, что более высокая концентрация цинка как и марганца находится в корнях, чем в листьях. А среди клеточных структур наиболее значительной концентрацией цинка отличаются вакуоли и незначительное содержание его обнаружено в клеточных стенках [138]. Способность же цинка образовывать лиганды с органическими кислотами частности цитрамона рассматривается [140] как основной механизм снижения его активности в вакуоли.
По данным [92] в листьях распределение цинка был следующим: 400 Мм в клетках эпидермиса, 100 Мм в мезофилле и 365 Мм в вакуолях. Согласно [142], содержание цинка в листьях варьирует от 0,66 до 1,29 мг/г. Более 60 % его находится в связанной с органическими веществами форме.
При изучении клеточного деления цинка в листьях гиперакумулятора Thlaspi caerulescens зависимости от содержания его в среде показано [92], что в клетках эпидермиса 6-суточных растений содержание цинка может достигать 400 Мм, а в мезофилле лишь 100 Мм. В вакуолях эпидермальных клеток концентрация цинка достигала 385 мм и была в 5 – 6,5 раз больше, чем в клетках мезофилла. Важно отметить, что содержание калия при этом снижался от 571 до 193 мм. Выявлено, что при наличии цинка в среде наблюдается угнетение активности К+ – каналов, т.е. цинк блокирует их функцию [83].
По данным [79] содержание цинка в зерне злаковых составляет 7,9-8,0 мг/кг. При применении же цинковых удобрений содержание цинка в семенах ячменя может возрастать от 10 до 20 и 30 мг/кг [90].
Показано чувствительность яровой пшеницы на предпосевную обработку семян. Внесение цинка увеличивает продуктивность растений [16]. Обработка семян пектином и микроэлементами значительно повышали энергию прорастания семян. Хелаты цинка и кобальта способствовали активизации роста и повышению урожая озимой пшеницы и сои на 4 – 5 ц/га [17]. Культуры, выращенные из семян с повышенным содержанием цинка дают высокие урожаи при выращивании на почвах с низкой обеспеченностью цинка [119]. Внесение органического удобрения и 25 кг/га ZnSO4 положительно влияет на урожай сельскохозяйственных культур [70]. Предпосевная обработка семян гречихи ZnSO4 в концентрации 0,1 – 0,05 % повышает устойчивость растений к наиболее вредоносным болезням [53].
Многочисленные исследования [79, 90] свидетельствуют о негативном влиянии дефицита цинка на процессы роста и урожай растений. Мгновенную реакцию растений на его дефицит (0,12 мкм против 77 мкм в питательной смеси) было установлено при изучении ростовых процессов кукурузы [125]. Угнетение роста стебля и других органов кукурузы было отмечено через 7 суток прорастания. Выявлено, что уменьшение концентрации цинка в органах растений негативно влияет на содержание ИУК. Кроме того, в цинк дефицитных растений томатов отмечено также снижение содержания ГК и АБК в листьях [135]. Согласно [136], цинк в культуральной среде ускоряет индуцированное ИУК корнеобразования.
В работах [107, 143] показано, что при дефиците цинка в меристематических клетках риса снижается содержание и активность Cu/Zn-супердиоксидисмутазы. Вместе с тем установлено [143], что концентрация белка Cu/Zn-супердиоксидисмутазы положительно коррелирует с устойчивостью генотипов пшеницы к дефициту цинка. При дефиците цинка наблюдалось [124, 127] снижение содержания хлорофилла и каротиноидов в листьях пшеницы и кукурузы, угнетение интенсивности фотосинтеза и активности карбоксилюючих ферментов и карбоангидразы [98, 153]. Согласно [125], оптимальным содержанием цинка в питательной среде является концентрация 0,5-0,8 мкм, а среди злаковых культур наиболее чувствительными к дефициту цинка являются растения твердой пшеницы [68, 115].
По данным [81] корни растений чувствительных к дефициту цинка (томаты и перец), характеризуются повышенной способностью к выделению корнями Zn-мобилизующих субстанций – сидератов необходимых для раствора водонерастворимых цинковых соединений: Zn3(PO4)2 и Zn(OH)2. При дефиците цинка нарастание массы томатов и перца подавлялось на 33-43 % против 15-25 % при выращивании люцерны и капусты. Но и при высоких концентрациях цинка в питательной смеси и увеличении его содержания в вегетативных органах наблюдается угнетение ростовых процессов и снижение урожая сельскохозяйственных культур. Исследованиями показано негативное влияние избыточного критического уровня цинка на активность транспорта ассимилятов. Согласно [56], избыток цинка к меди отрицательно действует на активность цитохром- и полифенолоксидазы. В цитоплазме довольно много цинка связана с белками, что обусловлено его способностью образовывать комплексы с органическими соединениями, в том числе органическими кислотами, аминокислотами, пуриновых и пиримидиновых оснований ДНК. Для выявления стабилизирующей и дестабилизирующей действия этого элемента на вторичную структуру ДНК были проведены исследования комплексообразования нативной и денатурированной ДНК с цинком. При этом установлено, что нативная ДНК образует комплексы в соотношении 1 эквивалент цинка на 1 нуклеотид, и выявлены специфичность его действия на структуру денатурированной ДНК. Показано, что связывание цинка с фосфатными группами и азотистыми основаниями зависит от состояния вторичной структуры ДНК и концентрации ионов цинка [9, 11]. Однако эти вопросы требуют дальнейшего исследования [12].
Для изучения роли цинка в ростовых процессах и азотном обмене растений были проведены исследования влияния его на активность ферментных систем синтеза ИУК, поглощения и компартментацию NO3- в структурах клеток корней и их энергетическое состояние. Исследованиями П.А. Власюка, К.Д. Каракис, Е.В. Рудаковой [13] показано, что при исключение цинка из питательной смеси активность ферментных систем биосинтеза ИУК ослабляется. На основании полученных данных сделан вывод о том, что цинк участвует в образовании триптофана на пути синтеза ИУК.
Таким образом, вопрос о зависимости между электрогенным транспортом Н+ ионов и ростом клеток требует дальнейшего исследования. Важным свойством каналов является избирательность. Оно заключается в пропускании определенных ионов и способности к изменению конформационного состояния, определяющим в среде их активность. Выполнение К+-каналами своих функций обусловлено особенностями их строения, включая селективный фильтр и датчик напряжения и другие структуры, обеспечивающие селективность и активность их действия.При рассмотрении литературных данных о кальциевых каналах отмечается, что кальций как основной элемент питания растений необходим для формирования структур, поддержки осмоса и осуществления сигнальных функций в клетках.
Анализ литературных данных свидетельствует о существенном влиянии марганца, бора, меди и цинка на активность ферментных систем, интенсивность фотосинтетических процессов, органогенез, и продуктивность растений. В меньшей степени изучено их действие на активность процессов, протекающих в корневой системе. Почти полностью отсутствуют данные об их воздействии на электрогенез и протон-движущей силы (ΔμН+) клеток корней.
Литература
1. Алексеева-Попова Н.В. Растения в стрессовых условиях минерального питания // Пробл. ботан. на рубеже 20-21 вв.: Тез. докл., представл. 2 (10) Съезд Рус. ботан. о-ва, Санкт-Петербург, 26-29 мая, 1998. Т. 1. – СПб, 1998. – С. 144.
2. Анспок П.И. Микроудобрения. – Л.: Агропромиздат 1990. – 272 с.
3. Барбер С.А. Биологическая доступность питательных веществ в почве. – М.: Агропромиздат, 1988. – 378 с.
4. Батов А.Ю., Максимов Г.В. Регуляция транспорта катионов через плазмолемму клеток корней кукурузы // Физиология и биохимия культ. растений. – 1995. – Т. 25, № 1. – С. 3 – 7.
5. Воробьев Л.Н., Егорова Н.Н., Рубин А.Б. Диагностика активности Н+- насосов растений как инструмент физиологической селекции // 2 Съезд Всес. о-ва физиологов раст., Минск, 24-29 сент. 1990 г: тезисы докл. – Ч. 1. – М., 1990. – С. 22.
6. Воробьев Л.Н. Регулирование ионного транспорта: теоретические и практические аспекты минерального питания растений. – М.: ВИНИТИ, 1988. – 161 с.
7. Вахмистров Д.Б., Мазель Ю.А. Поглощение и передвижение солей в клетках растений // Итоги науки и техники ВИНИТИ. Физиология растений. – М.: Наука, 1973. – Т.1. – С. 162 – 212.
8. Власюк П.А. Биологические элементы в жизнедеятельности растений. – К.: Наук. думка, 1969. – 516 с.
9. Власюк П.А. Марганцеве живлення і удобрення рослин. – К.: Вид-во УАСГН, 1962. – 421 с.
10. Власюк П.А. Микроэлементы и радиоактивные изотопы в питании растений. – К.: Из-во АН УССР, 1956. – 115 с.
11. Власюк П.А. Значение микроэлементов в нуклеиновом обмене растений // Физиология и биохимия культ. растений – 1971. – 3, № 3. – С. 276 – 286.
12. Власюк П.А., Жидков В.А., Компаниец М.И. и др. Влияние марганца на особенности вторичной структуры ДНК растений // Физиология и биохимия культ. растений – 1970. – 2, № 2 – С. 122 – 128.
13. Власюк П.А., Каракис К.Д., Рудакова Э.В. Влияние отдельных предшественников ИУК на ее ферментативный синтез в ранние периоды цинковой недостаточности у кукурузы // Физиология и биохимия культ. растений – 1973. – 5, № 6 – С. 13 – 19.
14. Власюк П.А., Климовицкая З.М. Физиологическое значение марганца для роста и развития растений. – М.: Колос, 1969. – 160 с.
15. Демидчик В.В., Соколик А.И., Юрин В.М. Поступление меди в растения и распределение в клетках, тканях и органах // Успехи соврем.биол. – 2001. – 121, № 2. – С. 190 – 197.
16. Дианова Т.Б. Влияние азота и микроэлементов на устойчивость яровой пшеницы к водным стрессам: Автореф. дис. канд. биол. наук. – М.: Моск. с.-х. акад., 1999. – 18 с.
17. Дозоров А., Исайчев В. Влияние хелатов и пектиновых веществ на посевные качества семян // Междунар. с.-х. ж. – 1998. – № 5 – С. 57 – 59.
18. Ивченко В.И, Ковальчук М.И. Поступление и физиологические функции молибдена в растениях // Микроэлементы: поступление, транспорт и физиологические ф-ции в растениях. – К.: Наук. думка , – 1987. – С. 109 – 161.
19. Иламанова Р.А. Влияние микроэлементов на содержание хлорофилла, углеводов и дыхание дынь // Изв. Ан ТССР Сер. биол. н. – 1991. – № 1. – С. 62 – 72.
20. Камбурова М., Рацков В., Гербанов Ст. Съдържание на манган в някои зеленчукови култури: [Докл.] Науч.-практ. конф. “Екол. пробл. на земл. ” Пловдив, дек., 1993 // Науч. тр. / Висш селскостоп. инст., Пловдив. – 1993. – 38, № 2. – С. 145 – 147.
21. Кибаленко А.П. Бор в жизни и продуктивности растений. – К.: Наук. думка, 1973. – 220 с.
22. Кириченко Е.Б., Чагвардиефф П., Мартынов О.Л., и др. Действие тяжелых металлов на высшие растения // Пробл. ботан. на рубеже 20-21 вв.: Тез. докл., представл. 2(10) Съезду Рус.ботан.о-ва, Санкт-Петербург, 26-29 мая, 1998. Т.1 – СПб, 1998. – С. 171.
23. Климовицкая З.М. Роль корневой системы в процессах поглощения и превращения марганца // Изд-во АН СССР, сер. биол. – 1961. – № 5. – С. 560 – 576.
24. Климовицкая З.М., Лобанова З.И., Прокопивнюк Л.М. Соотношение низкомолекулярной и высокомолекулярной РНК У растений при марганцевом хлорозе // Физиология и биохимия культ. растений . – 1973. – 5, № 6. – С. 131 – 135.
25. Ковалевич З.С. Влияние микроэлементов на содержание азотистых соединений и аминокислотный состав зерна гороха // Микроэлементы в биол. и их применение в с. х. и мед. : Тез. докл. 11 Всес. конф., Самарканд, 1990. – С. 292 – 293.
26. Конопля М.І., Москова Т.М. Мінеральне живлення та іонний гомеостаз рослин Луганськ, Альма-матер, 2001. – 135 с.
27. Мазель Ю.Я., Вахмистров Д.Б. Поглощение и передвижение солей в клетках растений // Итоги науки и техники, ВИНИТИ. Физиология растений. – М.: НАУКА, 1973. – Т. 1, № 5 – С. 164 – 212.
28. Медведев С.С., Батов А.Ю., Мошков, Маркова М.В. Роль ионных каналов у трансдукции ауксинового канала // Физиол. растений. – 1999. – т. 46, № 5. – С. 711 – 717.
29. Медведев С.С., Макарова И.В. Участие ионов кальция в процессах роста и морфогенеза // Физиология и биохимия культ. растений. – 1992. – Т. 24, № 3. – С. 281 – 286.
30. Медведев С.С., Маркова И.В., Катриченко М.И., Штонда И.А. Участие бора и кальция в гравитропической реакции колеоптилий кукурузы // Вестн. ЛГУ. – 1990. – Сер. 3, № 2. – С. 64 – 68.
31. Микроэлементы поступление транспорт и физиологические функции в растениях. – К.: Наук. думка, 1987. – 184 с.
32. Новак В.А., Иванкина Н.Г. Клеточный уровень АТФ, транспорт калия и электрические характеристики плазмалеммы элодеи при действии феррицианида // ДАН СССР. – 1986. – 286, №2. – С. 498 – 501.
33. Новак В.А., Миклашевич А.И. Феррицианидредуктазная активность листьев элодеи и ее связь с энергетическим метаболизмом. // Физиология растений. – 1984. – 31, №3. – С. 489 – 495.
34. Опритов В.А., Пятыгин С.С., Ретивин В.Г. Биоэлектрогенез у высших растений. – М.: Наука, 1991. – 216 с.
35. Оканенко А.С., Берштейн Б.Й. Калий, фотосинтез и фосфорный метаболизм у сахарной свеклы. – Киев: Наукова думка, 1969. – 212 с.
36. Островская Я.К. Физиологическая роль меди и основы применения медных удобрений. – К.: Изд-во УАСХИ, 1961. – 300 с.
37. Полевой В.В. Физиология растений. – М.: Высшая школа, 1989. – 464 с.
38. Полевой В.В., Саламатова Т.С. Физиология роста и развития растений. – Л.: Изд-во ЛГУ,1991. – 239 с.
39. Пузина Т.И. Значение гормонального баланса в реакции растений картофеля на условиях минерального питания // Агрохимия. – 2000. – № 4. – С. 27 – 32.
40. Пятыгин С.С. Распространяющиеся электрические сигналы в растениях // Цитология. – 2008. – 50, № 2. – С. 154 – 158.
41. Ринькис Г.Я., Рамане Х.К. Несбалансированность питания полевых культур макро- и микроэлементами в производственных условиях Латвийской ССР // Материалы ХІ Всесоюз. конф. “Микроэлементы в биологии и их применение в с.-х. и медицине”. – Самарканд, 1990. – С. 220 – 222.
42. Ринькис Г.Я., Ноллендорф Сбалансированное питание растений макро- и микроэлементами. – Рига: Зинатне, 1982. – 304 с.
43. Тарчевский И.А. Сигнальные системы клеток растений. – М.: Наука, 2002. – 294 с.
44. Тихонова Л.И. Ионные каналы вакуолярной мембраны высших растений // Биол. Мембраны. – 1998. – т. 15, № 3. – С. 245 – 258
45. Терещенко А.Ф. Функционирование протонной помпы и механизм действия ауксинов // Физиология и биохимия культ. растений. – 1994. – т. 26, № 1. – С. 3 – 11.
46. Терещенко А.Ф. ИУК-индуцированые изменения рН среды инкубации гипокотили созревающих семян фасоли. // Там же – 1992. – т. 24, № 4, – С. 372 – 375.
47. Титика М., Лисник С. Тома С. Активность и изоферментный состав нитратредуктазы у сахарной свеклы в зависимости от обеспеченности растений молибденом, бором и условий увлажнения // Ин-т физиол. раст. АН Респ. Молдова. – Кишинев, 1998. – 8 с.
48. Тома С.И., Лисник С.С., Великсар С.Г., Кауш М.В. Регулирование адаптативных реакций и продуктивности растений с помощью микроэлементов // Изв. АН Респ. Молдова. Биол. и хим. н. – 1991. – № 4. – С. 3 – 11.
49. Удобрения и препараты с микроэлементами. – К.: Наук. думка , 1975. – 200 с.
50. Физиологические основы питания растений. – К.: Наук. думка , 1971. – 334 с.
51. Хапова С.А., Пеплова Н.С. Влияние меди на содержание белков и углеводов у Fragania ananassa в зависимости от метода размножения // Соврем. пробл. естествозн.: биол. и хим.: Сб. тез. обл. науч. конф. студ., аспирантов и мол. Ученых. – Ярославль, 1999. – С. 24 – 25.
52. Чумаченко И.Н., Прошкин В.А., Войтович Н.В. Перспективы применения микроудобрений // Химия в с. х. – 1995. – № 6. – С. 22 – 26.
53. Шевчук В.К. Мікроелементи в підвищенні стійкості гречки до найбільш шкодочинних хвороб // Вісн. агр. наук. – 1996. – № 10. – С. 33 – 35.
54. Школьник М.Я. Микроэлементы в питание растений. Физиология сельскохозяйственных растений. – М.: Изд-во МГУ, 1967. – Т. 2. – С. 128 – 203.
55. Abutidze M., Mchedlishvili N., Pruidze G. The effect of metal ions on tea leaves β-glucosidase activity // Bull. Georg. Acad. sci. – 2000. – 161, № 3. – P. 525 – 526.
56. Agarwala Sirish Chandra, Nautiyal Bachendra Datt, Chaterjee Chitralekha, Nautiyal Nirmala Variations in copper and zinc supply influence growth and activities of some enzymes in maise // Soil Sci. and Plant Nutr. – 1995. – 41, № 2. – P. 329 – 335.
57. Arsova Antoaneta Effect of soil acidity neutralization and copper toxicity on maize productivity, cjpper uptake and biomass cation content // Bulg. J. Plant Physiol. – 1996. – 22, № 3 – 4. – Р. 56 – 65.
58. Asad A. Boron requirements for sunflower and wheat // J. Plant Nutr. – 2002. – 25, № 4. – P. 885 – 899.
59. Babalakova Nadejda, Traykova Daniela. Copper-induced cupric-and ferric-chelate reduction by intact barley roots // Bulg. J. Plant Physiol. – 2001. – 27, № 3 – 4. – Р. 93 – 103.
60. Babalakova N., Traykova D., Copper induced modifications of tonoplast permeability and proton pump ion sensitivity depending on cjncentranion of calcium in solutions of barley roots: Abstr. 11th Congress of the Federation of European Societies of Plant Physiologi, Varna, 7-11 Sept., 1998 // Bulg. J. Plant Physiol. – 1998. – Spec. issue. – Р. 120.
61. Barr R. The possible role of redox-associated protons in growth of plant cells // Journal of Bioenergetics and Biomembranes. – 1991. – 23, № 3. –P. 443 – 467.
62. Blaser-Grill Jürgen, Knoppik Dietmar, Amberger Anton, Goldbach Heiner. Influence of boron on the membrane potential in Elodea densa and Helianthus annuus roots and H+ extrusion of suspension cultured Daucus carota cells // Plant Physiol. – 1989. – 90, № 1. – Р. 280 – 284.
63. Blevins Dale G., Lukaszewski Krystyna M. Boron in plant structure and function // Annu. Rew. Plant Physiol. And Plant Mol. Biol. Vol. 49. – Palo Alto (Calif.), 1998. – Р. 481 – 500.
64. Blevins Dale G., Reinbott Tim M., Lukaszewski Krystyna M., Waters Brian M. Ironic problems in boron deficient, low ascorbate root tips: Abstr. Plant Biol. ’97: Annu. Meet. Amer. Soc. Plant Physiol., Vancouver, Aug. 2-6, 1997 // Plant Physiol. – 1997. – 114, № 3, Suppl. – P. 259.
65. Bowszys Teresa, Krauze Anna DziaŁanie nawozowe boru w uprawie buraka cukrowego // Acta Acad. agr. ac techn. olsten. Agr. – 1995. – № 61. – Р. 85 –91.
66. Buchanan B.B., Balmer Y. Redox regulation: a broadening horizon // Annu. Rev. Plant Biol. – 2005. – 56. – P. 187 – 220.
67. Bujtás Clara, Cseh Edith Connections between K+ and Cu2+ transport in winter wheat seedlings // 6th Inf. Trace Elem. Symp., Lepziq, 1989. Vol. 2 – Jena, 1989. – Р. 418 – 425.
68. Cakmak I., Torun B., Erenoğlu B., Öztürk L., Marschner H., Kalayci M., Ekiz H., Yilmaz A. Morphological and physiological differences in the response of cereals to zinc deficiency: Selec. Pap. 5th Int. Wheat Conf., Ankara, 6-10 June, 1996 // Euphytica. – 1998. – 100, № 1 – 3. – P. 349 – 357.
69. Camacho-Cristobal Juan j., Gonzalez-Fontes Agustin Boron deficiency causes a drastic decrease in nitrate content and nitrate reductase activity, and incrases th content of carbohydrates in leaves from tobacco plants // Planta. – 1999. – 209, № 4. – P. 528 – 536.
70. Сhannabasavanna A.S., Biradar D.P. Yield and yield attributes of transplanted summer rice as influenced by organic manures and zinc levels // J. Maharashtra Agr. Univ. – 2001. – 26, № 2. – P. 170 – 172.
71. Chatterjee Chitralekha, Nautiyal Nirmala. Varation in calcium levels leads to changes in the copper metabolism in Barley // Soil. Sci and Plant Nutr. – 2001. – 47, № 1. – Р. 9 – 16.
72. Colombo R., Cerana R., Lado P. Regulation of tonoplast K+channels by voltage in the range of physiological electric potentials // Plant Physiol. — 1990 — Vol. 93, № 1. – P. 350 – 352.
73. Dietrich P.. Dreyer 1., Wiesner P. et ai Cation sensitivity and kinetics of guard-cell potassium channels differ among species // Planta. — 1998. — Vol. 205, № 2. – P. 277 – 287.
74. Drew M.C., Webb J., Saker L.R. Regulation of K+ uptake and transport to the xylem in barley roots; K+ distribution determined by electron probe X-ray microanalysis of frozen-hydrated cells// J. Exp. Bot. – 1990. – Vol. 41, № 228. – P. 815-825.
75. Du C.W., Wang Y.H., Xu F.S., Yang Y.H., Wang H.Y. Study on the physiological mechanism of boron utilization efficiency in rape cultivars // J. Plant Nutr. – 2002. – 25, № 2. – Р. 231 – 244.
76. Fleischer Axel, Titel Christine, Ehwald Rudolf. The boron requirement and cell wall properties of growing and stationary suspension-cultured Chenopodium album L. cells // Plant Physiol. – 1998. – 117, № 4. – Р. 1401 – 1410.
77. Fleischer Axel, O’Neill Malcolm A., Ehwald Rudolf The pore size of non-graminaceous plant cell walls is rapidly decreased by borate ester cross-linking of the pectic polysaccharide rhamnogalacturonan II // Plant Physiol. – 1999. – 121, № 3. – Р. 829 – 838.
78. Fox G.G., Loughman B.C., McCallan N.R., Quigumpoix H., Ratcliffe R.G. NMR studies of manganese uptake and of manganese pretreated plants: Abstr. 9th Congr. Fed. Eur. Soc. Plant Physiol., Brno, 3-8 July, 1994 // Biol. plant. – 1994. – 36, Suppl. – P. 150.
79. Genc Yusuf, McDonald G.K., Graham R.D. Critical deficiency concentration of zinc in barley genotypes differing in zinc efficiency and its relation to growth responses // J. Plant Nutr. – 2002. – 25. № 3. – P. 545 – 560.
80. Goldbach Heiner E., Blaser-Grill Jürgen, Hartmann Dorothea, Porzelt Marion, Rötzer Thomas, Amberger Anton. Influence of B-deficiency on net proton-release of Daucus cells and membrane resistance of Elodea densa // Plant Physiol. – 1990. – 79, № 2, Pt 2. – Р. 93.
81. Hasegawa Isao, Ozawa Mariko, Noguchi Akira, Yazaki Jinya Relationship between the sensitivity to zinc deficiency of dicotyledous plants and the release of Zn- mobilizing substances from their roots: Pap. Annual Meeting and Symposia, Kyoto, March 28-30, 1999 // Plant and Cell Physiol. – 1999. – 40, Suppl. – P. 98.
82. Hino K., Kreij C. de, Elderen C.W. van Absorption of various types of chelated copper in a low concentration range by cucumber // J. Plant Nutr. – 1995. – 18, № 5. – Р. 1049 – 1056.
83. Hoth Stefan, Hedrich Rainer. Susceptibility of the guard-cell K+ - uptake channel KST1 to Zn2+ requires histidine redis ues in the S3-S4 linker and in the channel pore // Plant. – 1999. – 209, № 4. – Р. 543 – 546.
84. Hu Hening, Brown Patrick H., Labavitch John M. Species variability in boron requirement is correlated with cell wall pectin // J. Exp. Bot. – 1996. – 47, № 295. – Р. 227 – 232.
85. Ito O. Forms of boric acid in plant tissues and soil solutes observed with B-NMR // Trans. 14th Int. Congr. Soil Sci., Kyoto/ Aug., 1990. Vol. 4. Commis. 4- Kyoto, 1990. – Р. 718 – 719.
86. Johannes E., Brosnan J.M., Sanders D. Calcium channels and signal transaction in plant cells // Bioessays. – 1991. – Vol. 13, № 7. – P. 331 – 336.
87. Johannes E., Brosnan J.M., Sanders D. Calcium channels in the vacuolar membrane of plants: multiple pathways for intracellular calcium mobilization // Phil. Trans. Roy. Soc. London. B. – 1992. – Vol. 338, № 1283. – P. 105 – 112.
88. Kastori R., Plesničar M., Panković D., Sakač Z. Photosynthesis, chlorophyll fluorescence and soluble carbohydrates in sunflower leaves fs affected by boron deficiency // J. Plant Nutr. – 1995 – 18, № 9. – Р. 1751 – 1763.
89. Kennett E. C., Kuchel P.W. Redox reactions and electron transfer across the red cell membrane // IUBMB Life. – 2003. – 55. – P. 375 – 385.
90. Kenbaev Bakyt, Sade Bayram Response of field- grown baley cultivars grown on zinc-deficient soil to zinc application // Commun. Soil Sci. and Plant Anal. – 2002. – 33, № 3 – 4. – P. 533 – 544.
91. Kummerová M., Burešová I. The effect of manganese on biomass formation and the content of assimilation pigments in maize // Scr. fac. sci. nature. UJEP Brun. – 1989. – 19, № 1 – 2. – P. 63 – 70.
92. Küpper Hendric, Zhao Fang Jie, McGrath Steven P. Cellular compartmentation of zinc in leaves of the hyperaccumulator Thlaspi caerulescens // Plant Physiol. – 1999. – 119, № 1. – P. 305 – 311.
93. Lidon Fernando Cebola, Barreiro Maria da Graca, Henriques Fernando Santos Interactions between biomass production and ethylene biosynthesis in copper-treated rice // J. Plant Nutr. – 1995. – 18, № 6. – Р. 1301 – 1314.
94. Loomis W. David, Durst Robert W. Chemistry and biology of boron // Bio Factors. – 1992. – 3, № 4. – Р. 219 – 239.
95. Liu Peng, Yang Yu-ai Zhejiang daxue xuebao. Nongye yu shengming kexue ban Влияние стресса бора и молибдена на активность нитратредуктазы и содержание нитратного азота в листьях сои // J. Zhejiang Univ. Agr. and Life Sci. – 2000. – 26, № 2. – Р. 151 – 154.
96. Maathuis F., Ichida A., Sanders D. Roles of higher plant K+-channel // Plant
Physiol. – 1997. – 114, № 4. – P 1141 – 1149.
97. Macfie S.M., Cossing Ed. A., Taylor G.J. Effect of excess manganese on production of organic acids in Mn-tolerant and Mn-sensetive culti vars of Triticum aestivum L. (Wheat) // J. Plant Physiol. – 1994. – 143, № 2. – P. 135 – 144.
98. Macnair Mark R., Smirnoff Nick Use of zincon to study uptake and accumulation of zinc by zinc tolerant and hyperaccumulating plants // Commun. Soil Sci. and Plant Anal. – 1999. – 30, № 7 – 8. – P. 1127 – 1136.
99. Masarovičova E., Holubová M. Effekt of copper on growth and chlorophyll content of some herbs // Rostl. vyroba – 1998. – 44, № 6. – Р. 261 – 265.
100. Matoh Toru, Matsuda Akira, Kobayashi Masaru, Sekiya Hiro Localization of boron (B) in higher plant apoplast : Pap. Annual Meeting and Symposia, Kyoto, March 28-30, 1999 // Plant and Cell Physiol. – 1999. – 40, Suppl. – Р. 149.
101. Maywald Friedhelm, Weigel Hans Joachim Zur Biochemie und Molekularbiologie der Schwermetallaufnahme und-speicherung bei höheren Pflanzen // Landbauforsch. Volkenrode. – 1997. – 47, № 3. – Р. 103 – 126.
102. Mehlhorn Horst, Wenzel Andrea A. Manganese deficiency enhances ozone toxicity in bush beans (Phaseolus vulgaris L. cv. Saxa): [Pap] lst Int. Symp. Veg. Stress, Munich, June 19-21, 1995 // J. Plant Physiol. – 1996. – 148, № 1 – 2. – P. 155 – 159.
103. Mtoh Toru. Boron works in the cell wall: Abstr. Annu. Meet. and 37th Symp. Jap. Soc. Plant Physiol., March 27-29, 1997 // Plant and Cell Physiol. – 1997. – 38, Suppl. – Р. 5.
104. Mühling Karl Hermann, Wimmer Monika, Goldbach Heiner E. Apoplastic and membrane-associated Ca2+ in leaves and roots as affected by boron deficiency // Physiol. plant. – 1998. – 102, № 2. – P. 179 – 184.
105. Nautiyal N., Chatterjee C., Sharma C.P. Copper stress affects grain filling in rice // Comunn. Soil Sci. and Plant Anal. – 1999. – 30, № 11 – 12. – Р. 1625 – 1632.
106. Nowak Grzegorz A., Czapla Jerzy. Response of soybean to gibberellin A3 application under conditions of high boron availability // J. Plant Nutr. – 1995. – 18 № 10. – Р. 2179 – 2190.
107. Obata Hitoshi, Kawamura Satoko, Shimoyama Atsuo, Senoo Keishi, Tanaka Akiyoshi. Changes in the level of protein and activity of Cu/Zn-superoxide dismutase in zinc deficient rice plant, Oryza sativa L. // Soil Sci. and Plant Nutr. – 1999. – 45, № 4. – P. 891 – 896.
108. Ouzounidou G. Changes in variable chlorophyll fluorescence as a result of Cu-teatment: Dose-response relations in Silene and Thlapsi // Photosynthetica: – 1993. – 29, № 3. – Р. 455 – 462.
109. Ouzounidou Georgia, Moustakas Michael, Lannoye Robert Chlorophyll fluorescence and photoacoustic characteristics in relationship to changes in chlorophyll and Ca2+ content of a Cu-tolerant Silene compacta ecotype under Cu treatment // Physiol. plant. – 1995. – 93, № 3. – Р. 551 – 557.
110. Pádua M., Casimiro A. Maganese interaction on copper toxicity in pae chloroplast: Abstr. 9th Congr. Fed. Eur. Soc. Plant Physiol., Brno, 3-8 July, 1994 // Biol. plant. – 1994. – 36, Suppl. – Р.154.
111. Palavan- Ünsal N., Cetin E., Kadioglu A. Boron stress affects the peroxidase activity: Abstr. 11th Congress of the Federation of European Societies of Plant Physiology, Varna, 7-11 Sept., 1998 // Bulg. J. Plant Physiol. – 1998. – Spec. issue. – P. 266.
112. Pandey Nalini, Sharma C.P. Copper effect on photosynthesis and transpiration in safflower // Indian J. Exp. Biol. – 1996. – 34, № 8 – Р. 821 –822.
113. Pätsikkä Eija, Aro Eva-Mari, Tyystjärvi Esa Increase in the quantum yield of photoinhibition contributes to copper toxicity in vivo // Plant Physiol. – 1998. – 117, № 2. – Р. 619 – 627.
114. Pearson J.N., Rengel Z. Uptake and distribution of 65Zn and 54Mn in wheat grown at sufficient and deficient levels of Zn and Mn. I. During vegetative growth // J. Exp. Bot. – 1995. – 46, № 288. – P. 833 – 839.
115. Prasad K.V.S.K., Puthur Jos T., Sharmila P., Saradhi P.P. Detoxification of toxic oxygen species in crop plants exposed to zinc toxicity: Abstr. Plant Biol. ’97: Annu. Meet. Amer. Soc. Plant Physiol., Vancouver, Aug. 2-6, 1997 // Plant Physiol. – 1997. – 114, № 3, Suppl. – P. 56.
116. Rahman Hasan Hafuzur, Arima Yasuhiro, Watanabe Koichiro, Sekimoto Hitoshi Adequate range of boron nutrition is more restricted for root nodule development than for plant growth in young soybean plant // Soil. Sci. and Plant Nutr. – 1999. – 45, № 2. – Р. 287 – 296.
117. Ramirez R., Linares J.C. Boron deficiency symptoms and dry matter production of sesame under grennhouse conditions // Commun. Soil Sci. and Plant Anal. – 1995. – 26, № 17 – 18. – Р. 3043 – 3049.
118. Reinbott T.M., Blevins D.G., Schon M.K. Content of boron and branch leaves and seed of soybean // J. Plant Nutr. – 1997. – 20, № 7 – 8. – Р. 831–843.
119. Rengel Zdenko, Graham Robin D. Importance of seed Zn content for wheat growth on Zn-deficient soil // Plant and Soil. – 1995. – 173, № 2. – P. 267 –274.
120. Rizvi S. I., Jha R., Maurya R.K. Erytrocyte plasma membrane redox system in human aging // Rejuvenation Research. – 2006. – 9, № 4. – P. 470–474
121. Rubinstein B., Luster D. G. Plasma membrane redox activity: Components and role in plant processes // Annu. Rev. Plant. Physiol. Plant. Mol. Boil. – 1993. – 44. –P. 131 – 155.
122. Ruiz Juan M., Bretones German, Baghour Mourad, Ragala Lamia, Belakbir Amal, Romero Luis Relationship between boron and phenolic metabolism in Tobacco leaves // Phytochemistry. – 1998. – 48, № 2. – Р. 269 – 272.
123. Ruszkowska Maria, Łyszcz Stanisława, Faber Antoni, Filipiak Krystyna Prydatnosc niektorych wskaznikow zaopatrzenia pszenicy ozimej w miedz // Pam. Puław. – 1990. – № 96. – Р. 57 – 69.
124. Salama Z.A., Lazova G.N., Stoinova Zh.G., Popova L.P., El-Fouly M.M. Effect of zinc deficiency on photosynthesis in chick-pea and maize plants // Докл. Бълг. АН. – 2002. – 55, № 3. – Р. 65 – 68.
125. Sekimoto Hitoshi, Hoshi Misuo, Nomura Takahito, Yolota Takao Zinc deficiency affects the levels of endogenous gibberellins in Zea mays L. // Plant and Cell Physiol. – 1997. – 38, № 9. – P. 1087 – 1090.
126. Schon Mary K., Novacky Anton, Blewins Dale G. Boron induces hyperpolarization of sunflower root cell membranes and increases membrane permeability to K+ // Plant Physiol. – 1990. – 93, № 2. – Р. 566 – 571.
127. Sharma P.N., Bisht S.S., Kumar Praveen, Mishra M.K. Induction of oxidative stress by deficiency and toxicity of zinc in wheat plants grown in solution culture // Indian J. Agr. Biochem. – 1999. – 12, № 1. – P. 10 – 13.
128. Sharma P.N., Ramchandra Tanuja Water relations and photosynthesis in mustard plants subjected to boron deficiency // Indian J. Plant Physiol. – 1990. – 33, № 1. – Р. 150 – 154.
129. Shelp Barry J., Marentes Eduardo, Kitheka Alice M., Vevekanandan Pathmanathan. Boron mobility in plants // Physiol. plant. – 1995. – 94, № 2. – Р. 356 – 361.
130. Shen Zhenguo, Liang Yongchao, Shen Kang Effect of boron on the nitrate reductase activity in oilseed rape plants // J. Plant Nutr. – 1993. – 16, № 7. – P. 1229 – 1239.
131. Srivastava N.K., Luthra R. Influence of boron deficiency on 14CO2 and [U-14C] saccharose incorporation in primary metabolites in relation to essential oil accumulation in Mentha piperita // Photosynthetica. – 1993. – 29, № 3. – Р. 437 – 445.
132. Stancheva I., Kaloianova N., Atanasova E. Effect of copper and arsenic on the yield and plastid pigment content of rice inoculated with Azospirilum brasilerse // Почвозн., агрохим. и екол. – 1999. – 34, № 4 – 5. – Р. 140 – 144.
133. Taiz L, Zeiger E. Plant Physiology. – Sunderland Massachusetts: Sinauer Asso-ciates Inc. Publishers, 1998. – 791 p.
134. Tavakoli N., Kluge C., Golldack D., Mimure T., Dietz K.-J. Reversible redox control of plant vacuolar H+-ATPase activity related to disulfide bridge formation in subunit E, as well as subunit A // Plant J. – 2001. – 28, № 1. – P. 51 – 59
135. Takahashi Hisamitsu, Kamata Atsushi Влияние дефицита цинка на устьица листьев и содержание эндогенных гибберелловой и абсцизовой кислот в растениях томата (Lycopersicon eseulentum Mill) // Tokyo nogyo daigaku nogaku shuho=J. Agr. Sci. – 1997. – 42, № 1. – Р. 12 – 19.
136. Takaki Hiroshi, Matuo Hirofumi, Yasukouchi Ayako, Saeki Yuiichi, Nagatomo Yoshitaka. “Auksin – induced root formation” and zinc: Pap. Annual Meeting and Symposia, Kyoto, March 28-30, 1999 // Plant and Cell Physiol. – 1999. – 40, Suppl. – Р. 45.
137. Trewavas A. I. Growth substance sensitivity limiting factor of plant development // Physiol. рlant. – 1982. – 55, № 1. – P. 60 – 72.
138. Väzquez M.D., Barcelo J., Poschenrieder Ch., Madico J., Hatton P., Baker A.J.M., Cope G.H. Localization of zinc and cadmium: Thlaspi cearulescens (Brassicaceae), a metallophyte that can hyperaccumulate both metals // J. Plant Physiol. – 1992. – 140, № 3. – P. 350 – 355.
139. Ucoka Hanayo, Yoshiyuki Tanaka, Masayoshi Macshuma. Kinetic properties ol Ca2+-ATPaseandCa2+/H+antiporter of plant vacuolar membrane //Annu. Mccl. and 37th Symp. Jap. Soc. Plant Physiol., March 27-29, 1997: Abstr. – Plant and Cell Physiol. – 1997. – Vol. 38, Supll. – P. 136.
140. Wang Jian, Evangelou Bill P., Nielsen Mark T., Wagner George J. Computer, simulated evaluation of possible mechanisms for sequestering metal ion activity in plant vacuoles // Plant Physiol. – 1992. – 99, № 2. – P. 621 – 626.
141. Wang Z.Y., Tang Y.L., Zhang F.S., Wang H. Effect of boron and low temperature on membrane integrity of cucumber leaves // J. Plant Nutr. – 1999. – 22, № 3. – Р. 543 – 550.
142. Wu Yong-yao, Zhou Yi-feng, Zhou Da-zhai Hubei minzu xueyuan xuebao. Ziran Kexue ban Изучение распределения цинка в Brasenia schreberi // J. Hubei Inst. Nat. Natur Sci – 2001. – 19, № 4. – P. 1 – 3.
143. Yu Q., Worth C., Rengel Z. Using capillary electrophoresis to measure Cu/Zn superoxide dismutase concentration in leaves of wheat genotypes differing in tolerance to zing deficiency // Plant. Sci. – 1999. – 143, № 2. – Р. 231 – 239.
144. Yu Xiaohe, Belll Paul F. Nutrient deficienty symptoms and boron uptake mechanisms of rice // J. Plant Nutr. – 1998. – 21, № 10. – P. 2077 – 2088.